Earlier this year I was invited by the editor of British Wildlife magazine to write a piece for their Changing Perspectives section about how odd Europe is when it comes to bird pollination. It’s based on one of the chapters in my book Birds & Flowers: An Intimate 50 Million Year Relationship.
If you subscribe to the magazine, it will appear in the August issue, though I’m happy to send a PDF to anyone who doesn’t subscribe (or has not read the book) – use the Contact Page. The main accompanying photograph is by one of my former students, Lisa King, who kindly allowed me to use it.
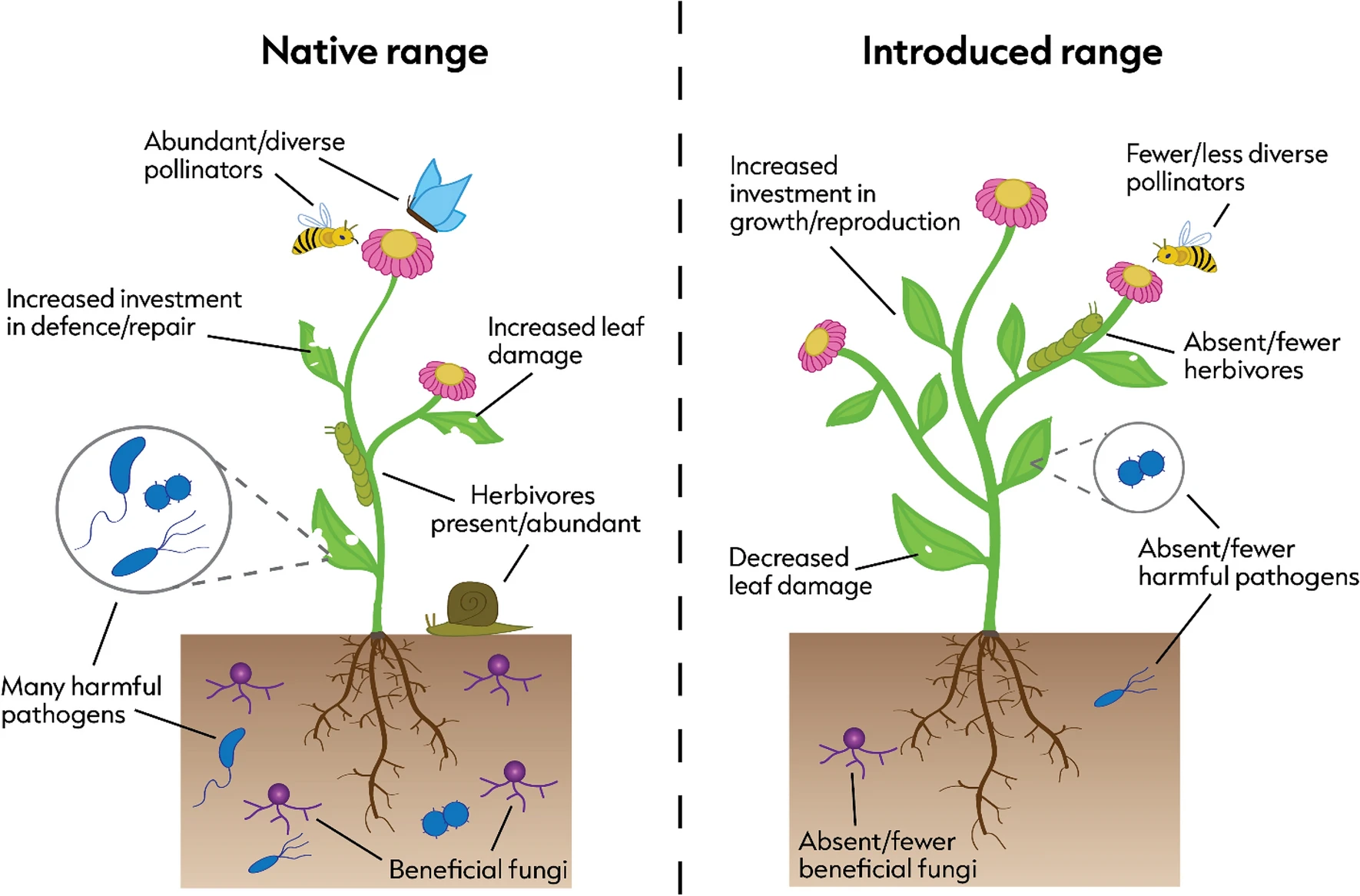
As I’ve previously discussed on the blog, when species are moved to a different part of the world they lose many of the ‘enemies’ – such as predators, herbivores and pathogens – that would normally keep their populations in check. This can have implications for the likelihood of a species becoming invasive, and it’s called the Enemy Release Hypothesis (ERH) and has been well studied. Less well researched is the flip side of the ERH, the Missed Mutualist Hypothesis (MMH), in which species lose their ‘friends’, such as pollinators, seed dispersers, symbiotic fungi, and so forth. It’s a topic I’ve worked on with my colleagues at the University of New South Wales, principally Angela Moles and her former PhD student Zoe Xirocostas.
Another paper from Zoe’s PhD work has just been published and in it she carried out a comparison of European plants that have been transported to Australia, and asked whether they had fewer pollinators in their new range. It turns out that they do!
Here’s the full reference with a link to the paper, which is open access:
Many studies seeking to understand the success of biological invasions focus on species’ escape from negative interactions, such as damage from herbivores, pathogens, or predators in their introduced range (enemy release). However, much less work has been done to assess the possibility that introduced species might shed mutualists such as pollinators, seed dispersers, and mycorrhizae when they are transported to a new range. We ran a cross-continental field study and found that plants were being visited by 2.6 times more potential pollinators with 1.8 times greater richness in their native range than in their introduced range. Understanding both the positive and negative consequences of introduction to a new range can help us predict, monitor, and manage future invasion events.
Is it too early to talk about Christmas? Not if you’re interested in pollinators and pollination! The mid-winter festival has featured quite a number of times on my blog over the years, especially in relation to the iconic plants that represent this time of year in Northern Europe, and what one might describe as the ‘cultural biodiversity‘ of Christmas. The final plant that I included in that last post was the poinsettia (Euphorbia pulcherrima) – this is how I described it:
In many ways this is an unusual plant to have such a strong cultural association with Christmas: it’s a mildly toxic species of spurge from tropical Mexico that was introduced to North America in the 19th century, then subsequently to Europe. However its festive connotations date back to the earliest period of Spanish colonisation in the 16th century, so it’s older than some…other Christmasy traditions…
Just occasionally one sees a bird-pollinated tree planted in a city. The most common in my experience are various banksias in Australia, and the Royal Poinciana (from Madagascar) and the African Tulip Tree in the urban tropics and subtropics elsewhere in the world. I’ve also occasionally encountered large specimens of Poinsettia: when they are given free rein they are a much more impressive plant than their Christmas cousins. The vivid red bracts that surround the clusters of flowers suggest that they may be hummingbird-pollinated in their native Central America, but as far as I know their pollination ecology has not been studied.
Here at the Kunming Botanic Garden there’s several quite large specimens of poinsettia that, as I write, are in full flower, their red bracts a signal to pollinators that can be seen for quite a distance. However we’ve not seen any of the local sunbirds or white-eyes visit the flowers, and, as I said in the book, as far as I know the pollination ecology of poinsettia has never been studied in the wild. Close inspection of the flowers in the garden revealed that almost all of the nectaries had at least one nectar-collecting ant sticking out from it, their prominent backsides a deterrent to the Asian Honey Bees (Apis cerana) that also wanted a piece of the action.
Based on the position of the nectaries in relation to the stamens, if the plant is hummingbird-pollinated then the pollen is likely to end up under the chin of the bird. That’s certainly been described in other plant-bird pollination systems. But it does not have to be birds that move the pollen around – red flowers are also associated with other kinds of pollinators, for example butterflies and beetles. But until someone in Mexico does the necessary field work, we’ll just have to speculate.
The reintroduction of the Chequered Skipper butterfly to England is one of the outstanding conservation success stories of the last ten years. I’ve been proud to play a part – see these old posts here, here and here – and in particular supervising Jamie Wildman’s PhD work. The second paper from his thesis has just been published and in it Jamie documents how you can identify individual butterflies by their markings and use this information to estimate the population size, life-span, and movements of Chequered Skippers. The technique could also be applied to other distinctively marked butterflies.
Here’s the reference with a link to a read-only version of the study:
If you need a PDF, get in touch via my Contact page.
Here’s the abstract:
The chequered skipper butterfly Carterocephalus palaemon was reintroduced to Fineshade Wood, England in 2018 as part of a Butterfly Conservation-led project following several years of planning. From 2019–2022, the population was sampled each May–June by the lead author, timed count volunteers, Butterfly Conservation staff, and casual observers.
A novel photographic mark-recapture (PMR) technique was trialled as an alternative to mark-release-recapture (MRR). In conjunction with timed counts, PMR was used to photoidentify individual C. palaemon through each butterfly’s upperside (ups) wing markings, estimate daily and gross population size, detect movements, and determine lifespan. As capture and recapture can be achieved non-invasively using PMR, habitat disturbance, the potential to influence butterfly behaviour, accelerate wing wear, affect mate selection and predation, and heighten mortality risk through handling are eliminated. We found PMR to be a viable alternative to MRR for a sensitive reintroduction of a low-density species with unique ups markings such as C. palaemon. Using capture histories generated through PMR, from a known founder population size of 42 butterflies in 2018, we estimated the population at Fineshade Wood had increased to 618 butterflies (+ 1371.43%) by 2022.
Movements of up to 2.22 km over a time period of 17 days were also detected. Lastly, we discuss the implications of PMR for population sampling of other Lepidopterans, and the potential to improve cost-efficiency of the technique using machine-based learning tools.
Back in August 2022, Karin and I traveled to Kenya where I was teaching on a Tropical Biology Association field course at the Mpala Research Centre – see my posts from the time here and here.
Students on the course have to complete an extended group project, with supervision by teaching staff. Two of the groups looked at the visitors to flower heads of one of the dominant savannah acacias and the interactions between wild honey bees of the native subspecies and the other insects. There have been rather few studies of this honey bee in the wild and so we wrote up the work as a short research note that has now been published in the African Journal of Ecology.
The photo above shows the authors – ‘Team Etbaica’ – from left to right: Luis Pfeifer, Swithin Kashulwe, me, Caka Karlsson, and Janeth Mngulwi.
Here’s the reference with a link to the publisher’s site – the paper is open access:
The East African lowland honey bee (Apis mellifera scutellata) is reported as an aggressive subspecies of the Western honey bee, but few studies have investigated the impact of its aggressiveness on other insect pollinators. Observations of flower visitors to Vachellia (Acacia) etbaica and interactions between honey bees and other insects were conducted in 2022 in Mpala, Kenya. A total of 873 individual flower visitors were recorded, the most frequent being Hymenoptera, followed by Diptera and Lepidoptera. Honey bees dominated floral resources in the morning and late afternoon. When honey bees encountered other types of insects, they displaced the latter from flowers 100% of the time. This has never been observed in other Western honey bee subspecies, and we recommend further research on these taxa.
During the 2020 lockdown caused by the COVID-19 pandemic, I coordinated an international network of pollination ecologists who used standardised methods to collect data in their gardens. I blogged about it at the time – see here and here for instance – and also put up a post when the data paper from that work was published.
Several research groups are now working with that huge data set and interrogating it for answers to a wide range of questions. The first group to actually publish a paper from the data is a largely Chinese set of researchers from the Key Laboratory of Plant Resources, Conservation and Sustainable Utilization, at the South China Botanical Garden in Guangzhou, assisted by Kit Prendergast and myself.
In this paper we’ve considered how robust these plant-pollinator networks are to simulated extinctions of species, and how this is affected by the elevation, latitude, and plant species diversity of the network.
Here’s the full reference with a link to the study:
If you can’t access it and need a PDF, please send me a request via my Contact page.
Here’s the abstract:
Plant-pollinator interactions play a vital role in the maintenance of biodiversity and ecosystem function. Geographical variation in environmental factors can influence the diversity of pollinators and thus, affect the structure of pollination networks. Given the current global climate change, understanding the variation of pollination network structure along environmental gradients is vital to predict how global change will affect the ecological interaction processes. Here, we used a global plant-pollinator interaction data collection by the same sampling method at the same period to explore the effects of elevation, latitude, and plant richness on the structure and robustness of pollination networks. We analyzed a total of 87 networks of plant-pollinator interactions on 47 sites from 14 countries. We conducted a piecewise structural equation model to examine the direct and indirect effects of elevation, latitude, and plant richness on the network robustness and analyzed the function of network structure in elucidating the relationship between robustness and these gradients. We found that plant richness had both positive effects on robustness under random and specialist-first scenarios. Elevation, latitude, and plant richness affected network connectance and modularity, and ultimately affected network robustness which were mediated by nestedness under specialist-first and random scenarios, and by connectance under the generalist-first scenario. This study reveals the indirect effects of elevation, latitude, and plant richness on pollination network robustness were mediated by nestedness or connectance depended on the order of species extinctions, implying that communities with different pollination network structures can resist different extinction scenarios.
Researchers at the University of East Anglia (UEA) are calling for volunteers to contribute to their investigation of the springtime pollen diets of bumblebees in the UK countryside.
Spring is a critical season for colony-establishing bumblebees, but little is known about what types of pollen they collect at this time. In particular, trees, shrubs, and woodland flowers may make significant contributions to bumblebee pollen diets.
Volunteers are needed from across the UK to collect pollen samples from live bumblebees in April and May this year. These will then be analysed by the researchers to determine their taxonomic composition.
Volunteers can collect as many or as few samples as they like – every contribution will help! For an information pack detailing what is involved, and to receive a registration form, please email Guthrie Allen (BIO – Postgraduate Researcher): G.Allen [at] uea.ac.uk.
In my new book Birds & Flowers: An Intimate 50 Million Year Relationship I spend a bit of time discussing the idea of the bird pollination syndrome that we refer to as ‘ornithophily’, its limitations, and the fact that it has two distinct meanings that are often conflated. One of the problems with ornithophily, and indeed all of the syndromes, is that historically it’s sometimes blinkered scientists to the extent that they only look at the flower visitors that are “right” for the syndrome, ignoring the rest or dismissing them as “secondary pollinators”, a term I dislike.
Why do I dislike that term? Because it fails to capture the complexity of flower-pollinator interactions and relegates an important component of plant reproduction to a subsidiary role. I could go on about this at some length, but if you’re interested in discovering more, look at pages 62-65 of Birds & Flowers. There I contrast the classical Most Effective Pollinator Principle with the equally valid (but much less well studied) Least Effective Pollinator Principle, with a segue into one of my favourite tracks from Led Zeppelin’s second album: What is and What Should Never Be.
But back to the real subject of this post – a flower that corresponds to the classical bird pollination syndrome BUT is also pollinated by bees and (very surprisingly) wind! It’s such an interesting paper by Brazilian ecologists Amanda Pacheco, Pedro Bergamo & Leandro Freitas – here’s the reference and a link to the study:
For over 100 years the classical pollination syndromes have acted as a framework for understanding the ecology and evolution of plant-pollinator interactions. But we’ve long known that while they can be a useful shorthand, they do not fully reflect the complexity of how pollination systems evolve. That shouldn’t surprise us because, as I point out in my two recent books, we have data (of any quality) on no more than 10% of the 350,000 or so species of flowering plants!
In addition, those plants for which we do have good data are NOT a random subset of the flowering plants: they have been specifically chosen by researchers because they look to be good systems with which to address particular ecological or evolutionary questions.
Which is fine, but we MUST recognise that this imposes significant restrictions on our understanding of the biodiversity of plant-pollinator interactions. The authors of this paper expressed it very well when they wrote that assumptions about:
“predictability may cause researchers to take for granted that only birds pollinate ornithophilous flowers, hindering research on the contribution of other vectors.”
To which I’d add: it also hinders our understanding of how these interactions evolve over long time scales and across multiple populations.
An obvious question is: how frequent are these sorts of complex pollination systems, involving different pollen vectors of an apparently specialised flower? The answer is that we simply don’t know, because most researchers would have not gone into this level of detail. So a huge congratulations to the authors for a great study – I hope it stimulates others to look beyond the ‘expected’ pollinators of flowers.
Photos: Nathália Susin Streher from the original paper.
The final deliverable from the WorldFAIR Project with which I’m involved has recently been published and can be freely downloaded from Zenodo by following the link below. The report is called “Agricultural biodiversity FAIR data assessment rubrics” and in it we present the results from a series of six pilot studies that adopted the FAIR* standards and our recommendations from the previous report.
This document complements the previous one by giving examples and setting out guidelines that allow researchers and practitioners to ensure FAIRness in their plant-pollinator interaction data.
Here’s the full reference:
Drucker, D. P., Salim, J. A., Poelen, J., Soares, F. M., Gonzalez-Vaquero, R. A., Devoto, M., Ollerton, J., Kasina, M., Carvalheiro, L. G., Bergamo, P. J., Alves, D. A., Varassin, I., Tinoco, F. C., Rünzel, M., Robinson, D., Cardona-Duque, J., Idárraga, M., Agudelo-Zapata, M. C., Marentes Herrera, E., Taliga, C., Parr, C.S., Cox-Foster, D., Hill, E., Maués, M.M. Agostini, K. Rech, A.R., Saraiva, A. (2024). WorldFAIR (D10.3) Agricultural biodiversity FAIR data assessment rubrics (Version 1). Zenodo. https://doi.org/10.5281/zenodo.10719265
Although this is the last formal deliverable from our WorldFAIR work package, it’s not the final output that we have planned. I’ll report back on the journal paper(s) that we are writing as and when they are published.
*Findable, Accessible, Interoperable & Reusable (or sometimes Reproducible)
One of the projects in which I’m currently involved is the WorldFAIR project. Funded by the European Commission, WorldFAIR is exploring how to make data FAIR – Findable, Accessible, Interoperable and Reusable – across a range of different disciplines in the sciences and humanities.
My involvement is specifically with Work Package 10, which is focused on data standards for plant-pollinator interactions, particularly as they relate to pollination of agricultural crops.
I’m delighted to say that the second deliverable from that work – a set of standards, best practices and guidelines recommendations – is now available for free download if you follow the link in this reference:
Drucker, D., Salim, J. A., Poelen, J., Soares, F. M., Gonzalez-Vaquero, R. A., Ollerton, J., Devoto, M., Rünzel, M., Robinson, D., Kasina, M., Taliga, C., Parr, C., Cox-Foster, D., Hill, E., Maues, M. M., Saraiva, A. M., Agostini, K., Carvalheiro, L. G., Bergamo, P., … Trekels, M. (2024) WorldFAIR (D10.2) Agricultural Biodiversity Standards, Best Practices and Guidelines Recommendations (Version 1). Zenodo. https://doi.org/10.5281/zenodo.10666593
Here’s the first paragraph of the summary:
The WorldFAIR Case Study on Agricultural Biodiversity (WP10) addresses the challenges of advancing interoperability and mobilising plant-pollinator interactions data for reuse. Previous efforts, reported in Deliverable 10.1 – from our discovery phase – provided an overview of projects, best practices, tools, and examples for creating, managing and sharing data related to plant-pollinator interactions, along with a work plan for conducting pilot studies. The current report presents the results from the pilot phase of the Case Study, which involved six pilot studies adopting standards and recommendations from the discovery phase. The pilots enabled the handling of concrete examples and the generation of reusable materials tailored to this domain, as well as providing better estimates for the overall costs of adoption for future projects.