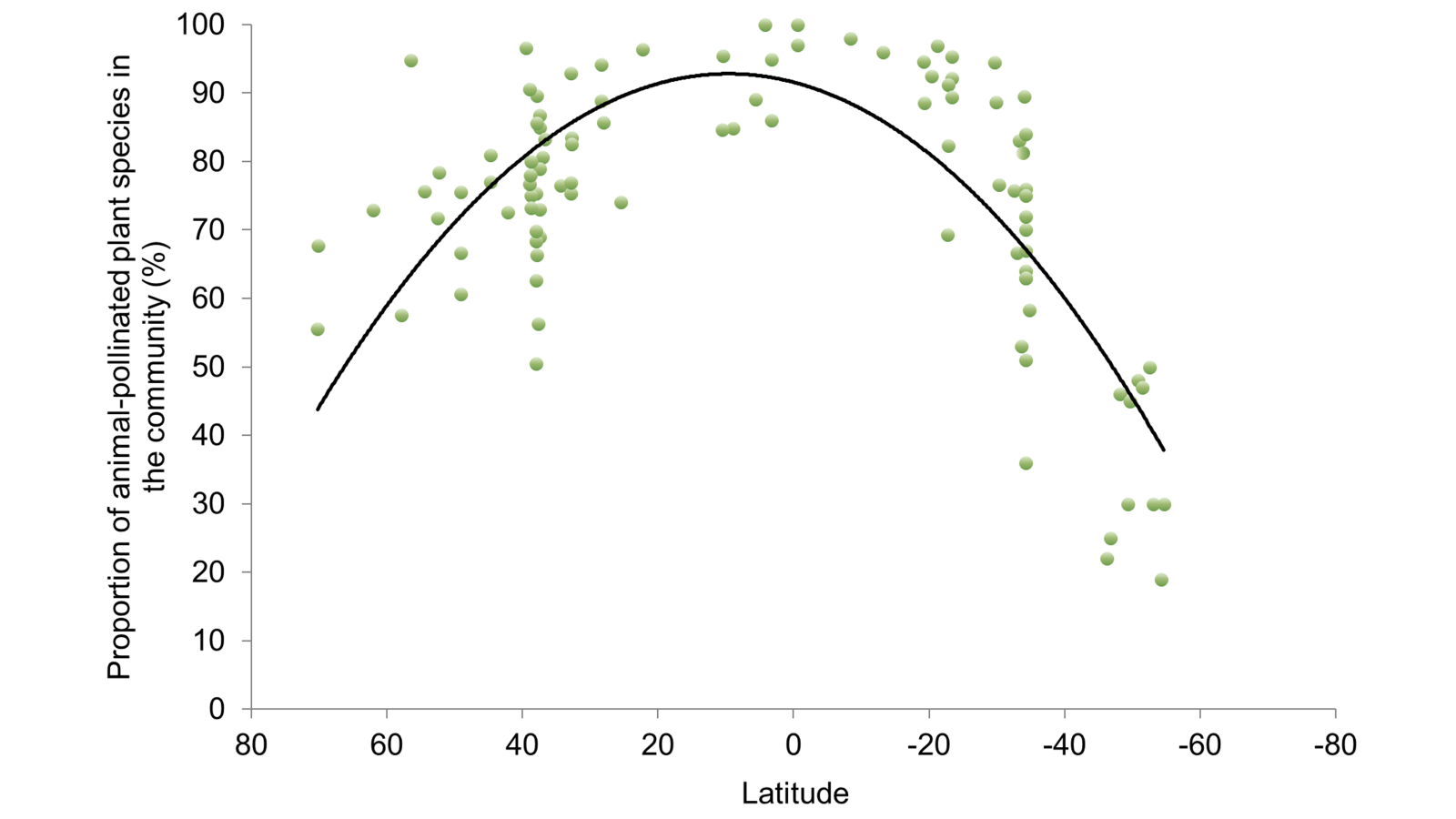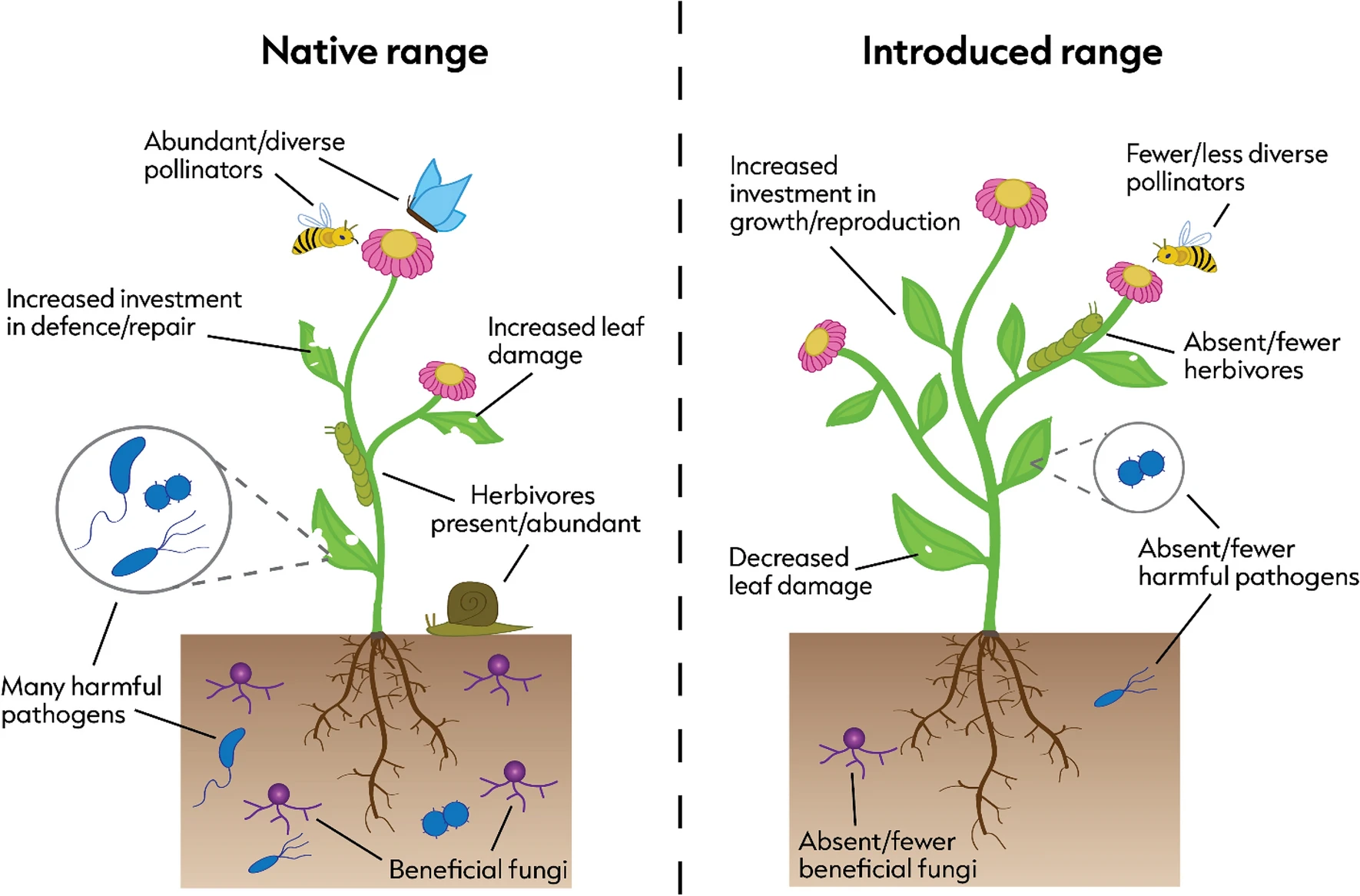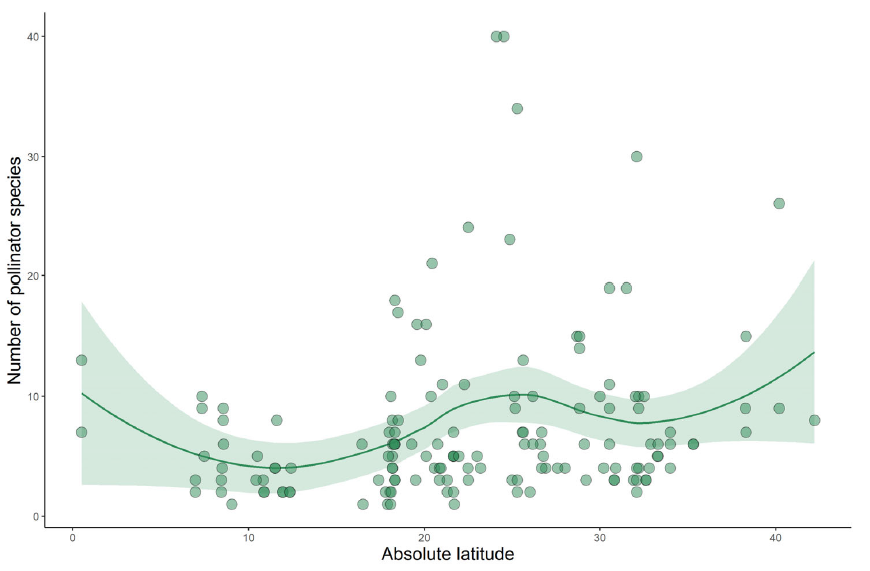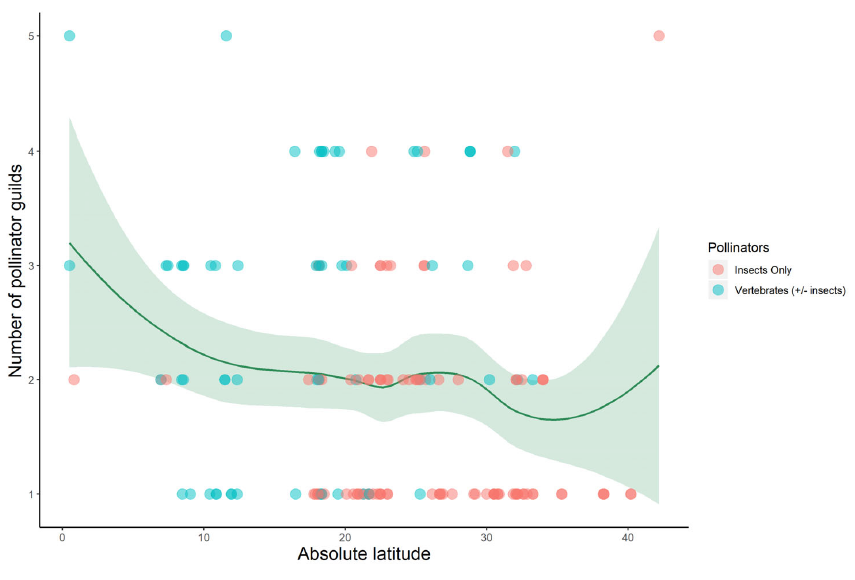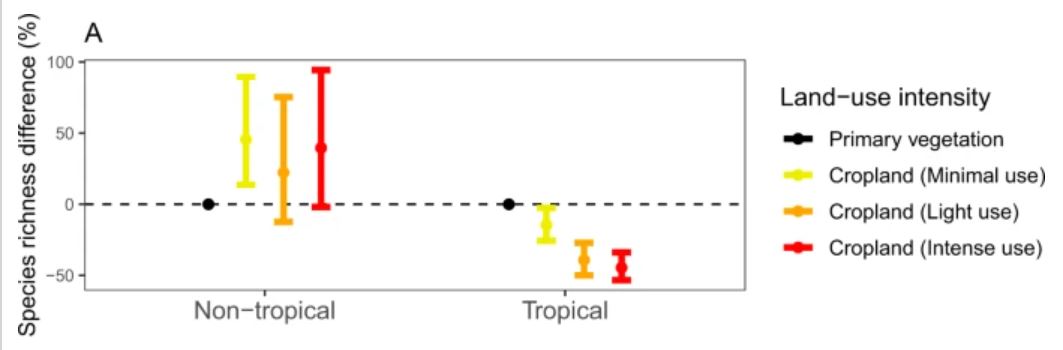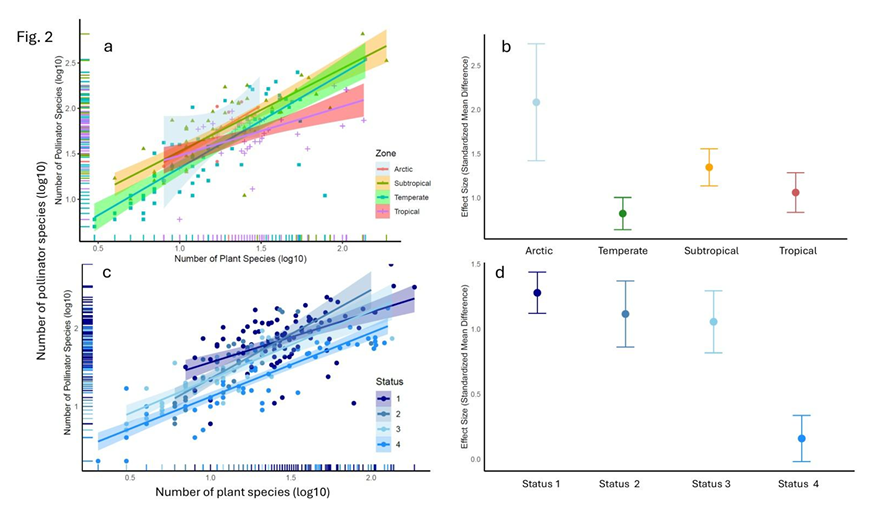
Last week I posted a preprint on the platform Research Square of a new manuscript entitled “The global relationship between flowering plant and pollinator diversity holds true across scales, latitude, and human influence” – follow that link to access a copy. The study is a collaboration with more than thirty colleagues and it develops some ideas that have been chugging around in my head for a number of years. It’s been reviewed and we’re at the stage of undertaking the revisions. I’m very excited to see it out in one form or another!
As far as I can recall this is the first time that I’ve been the lead author on a study that’s been posted as a preprint and I was not prepared for what happened after it went live on 2nd March!
Since then I’ve received over 30 invitations from journals to submit the paper for publication. Obviously, most (all?) of these are automated, because the majority are for journals that are in no way suitable, e.g., Insights of Herbal Medicine, Biomedical Science and Clinical Research, and my particular favourite, the Journal of Surgery Care!
I expected one or two spammy invitations like this, but not so quickly: the preprint went live at about 07:00 and the first request was received less than two hours later. Even now they are coming in at a rate of two a day.
It’s fairly clear that preprint servers are now being automatically mined by journal marketing algorithms. Within hours of a manuscript appearing online, the title, keywords, and author details are harvested and fed into bulk invitation systems. Can legitimate preprint publishers like Research Square not do anything about it?
Each email requires power to get it from a server to my Inbox, so as well as being irritating it’s a waste of resources. Presumably this strategy by these predatory publishers occasionally works with naive authors, otherwise they wouldn’t bother doing it. I’m almost (almost!) tempted to respond to one of these invitations and see what happens. But life’s too short.
Preprints are meant to accelerate open science and transparent peer review. Ironically, the same openness also makes it trivial for automated systems to harvest new manuscripts and generate waves of journal solicitations. None of this detracts from the value of preprints—they are a powerful way to share research quickly and openly—but it’s a reminder that openness in science inevitably attracts a few opportunists as well.
Anyway, if you’re planning to submit a preprint, don’t say that you weren’t warned – you may discover that a remarkable number of journals are suddenly desperate to publish your “valuable manuscript”.