Recently the Bumblebee Conservation Trust (BCT) reported that, in 2024, British bumblebees experienced their worst year since the BCT started its monitoring campaign. Overall, the numbers of bees were down by more than one fifth, with one of our commonest species, the Red-tailed Bumblebee (Bombus lapidarius) declining a staggering 74%! The cause seems to be the cold, wet spring of 2024 and we have to hope that this is a blip that will not be repeated in 2025. So far the year has been cold and I didn’t see my first queen bumblebee flying until early March. But the very warm weather over the last few days has encouraged bumblebees out of hibernation and plants to start flowering.
Long-term monitoring of the type that the BCT undertakes with its volunteers, is vital if we are to understand how British pollinators are faring. When I compiled the evidence for the chapter entitled ‘The shifting fates of pollinators’ in my book Pollinators & Pollination: Nature and Society, I tried to give a global overview, but also focused on British records, which are probably the best long-term data that is available on trends in pollinators. This information is compiled by the Joint Nature Conservation Committee (JNCC) as part of its annual UK Biodiversity Indicators reports. Each year it produces an indicator showing trends in bees, hoverflies, and the two combined as an overall pollinator trend*. To quote the JNCC website:
The indicator is based on 394 species (158 species of bee and 236 species of hoverfly), and measures change in the number of 1 kilometre grid squares across the UK in which they were recorded in any given year: this is referred to as the ‘occupancy index’.
The bee data comes from the Bees, Wasps and Ants Recording Society (BWARS) and the graph of bee trends that I used in that chapter of my book assessed records up until 2017. It looked like this:

As you can see, the index fluctuated a bit but was on average fairly stable up until 2005, after which there was a sharp decline, then an uptick from about 2014, though still low compared to the 1980 baseline. The overall impression is that bees had a tough time from the early 2000s onward, but things seem to be improving.
Since my book came out in 2021 I’ve given a lot of talks to natural history societies, ran training with consultancies and local councils, and so forth. Each year I update the JNCC graphs in my talks to give the audience the latest information. This is the one I used last year, which took the data up to 2019**:

This looks a bit different – the fluctuations are more pronounced – but overall the trend is similar, though the drop after 2015 is worrying. The impression is that there’s been big (cyclical?) fluctuations in the bee index over time, but its generally always below the 1980 baseline.
Updating the story to 2022 (the most recent available) shows a very different picture:

The impression it gives is that there’s been some modest fluctuations in the bee index, but then from about 2013 onward, the index has massively improved and now wild bees are doing better than ever!
What’s happening here? Why are these three graphs – published over a period of about five years – giving such different impressions of what’s happening to wild bees in Britain? As far as I can tell there’s two main reasons for the changes. The first is that the number of bee species included in the index increased from 137 to 148 to 158. Adding species for which there was previously no or little data is clearly going to have an effect.
The second reason, perhaps more fundamental, is that the method used for calculating the index has been refined, as explained in the technical annex to the study. That’s important because the data underlying the bee index was never collected in a standardised way for the purposes of assessing species’ trends. For this reason the UK Pollinator Monitoring Scheme (PoMS) was developed and it’s interesting to see that the data in the latest PoMS report shows some stability in wild bee abundance from 2017 to 2022:

So the latest data suggests that, for once, there’s some good news in the world of British wildlife. Does this mean that we should be complacent about the state of our wild bees? Absolutely not! As always, the devil’s in the details. The BCT report that I cited at the start of this post provides one level of (worrying) detail. But another is provided by the JNCC’s own statistics. As well as showing the overall trend in the bee index, the analysis digs into what is happening for individual species and provides a helpful summary figure like this:

Clearly many species are doing well, or at least have not changed since the 1980s. But more than one quarter of British wild bees are showing a weak or strong decline over the long term. That’s a clear signal that we need to keep on with our efforts to support wildlife and enhance our strategies to improve the state of nature in Britain.
As always, feel free to comment on the post or get in touch via my Contact page.
My sincere thanks to all of the volunteer naturalists who collect the data used by JNCC and PoMS – the task of assessing trends in wildlife would be impossible without your commitment!
————————————————————————————————–
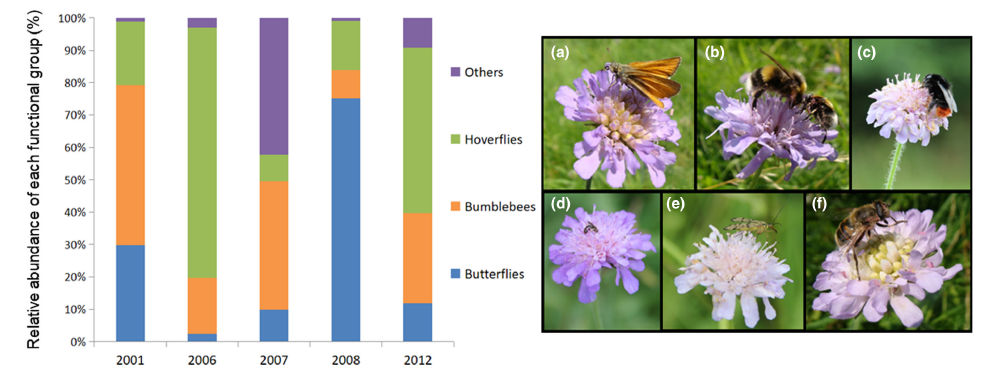
*Why JNCC does not include butterflies – which are assessed separately – in this overall trend is unclear to me, as we know that they can be important pollinators for some plants – see my blog post: ‘Butterflies, bumblebees and hoverflies can be equally effective pollinators of some plants says a new study‘.
**The data in the JNCC report is always a couple of years behind the publication date.