Earlier this week, the East Midlands Environment Agency proudly tweeted that they had placed honey bee hives on an ecologically important site that they own. As you might imagine, the response from pollinator experts such as myself, conservation NGOs, and some beekeepers, was not positive, as you can see if you look at the comments beneath my tweet:
If this is an important site for nature conservation, as claimed, then the #EnvironmentAgency should NOT put hives of honey #bees on it!
— Prof. Jeff Ollerton – @JeffOllerton@ecoevo.social (@JeffOllerton) July 12, 2023
By coincidence, overnight I received a message from someone in the USA asking for advice. Here’s a redacted version of their message:
My community has a 4 acre serpentine barren site that is part of a larger string of these unique barrens ….. Honey bee hives have recently been located adjacent to the barrens. Can you advise me as to the best way to determine whether there are, and to document any, adverse effects to the serpentine barrens native pollinators?
Going back to the question of how to assess any impacts, the simple answer is that it’s not easy and it relies on having good data. This was my response to my American correspondent:
Ideally you would need to take a before-and-after approach where you have data on things like number of native pollinator species, their abundance (including nest sites), rates of visitation of different pollinators to flowers, and fruit or seed set from particular plants. You’d then compare what was going on before the hives arrived with what’s occurring since their arrival.
If you don’t have the “before” data it’s much more difficult to assess if there has been an impact from the honey bees. However, the advice of most conservation groups is to adopt the “precautionary principle” and not site hives on or adjacent to areas of nature conservation value, especially if they are relatively small areas. See for example the Bumblebee Conservation Trust’s advice: https://www.bumblebeeconservation.org/managed-honeybees/
The precautionary principle is a well established concept across a range of areas, including health and engineering, as well as nature conservation. In the latter it needs to be more widely applied, especially when it comes to questions of where to site honey bee hives, and how many.
Last year I posted about the work that I’ve been doing on railways and biodiversity with UIC – the International Union of Railways – and the UK Centre for Ecology & Hydrology. That work is now complete and the second of our two reports called UIC Guidelines on Managing Railway Assets for Biodiversity is now out.
UKCEH has produced a press release and I’m copying it verbatim below:
New guidelines for the management of Europe’s railway network to protect and enhance biodiversity have been published.
The UK Centre for Ecology & Hydrology (UKCEH) has worked with the International Union of Railways (UIC), the professional association representing rail companies across the world, to provide technical recommendations and key design features for incorporating and enhancing habitats within the existing European network and new line upgrades, providing examples of best practice.
The UIC European region comprises 118-member companies from 39 countries, amounting to 350,000 kilometres of rail network.
Professor Richard Pywell, Head of Biodiversity at UKCEH who is one of the lead authors of the report, said: “We worked closely with railway companies across Europe to distil the best available knowledge on managing railway assets to benefit nature. For each asset, we considered the most effective measures to protect and restore biodiversity, and how to monitor the outcomes of these interventions.”
Another report author, independent consultant Professor Jeff Ollerton, added: “Working with UIC on this project has revealed just how important the land managed by Europe’s railway companies is for nature. The next step is to better understand how nature supports Europe’s economy, and the health and wellbeing of its people.”
The authors used the widely-adopted mitigation hierarchy approach which guides, developers on protecting existing habitats and ecosystems where possible.
The new UIC Guidelines for Managing Railway Assets for Biodiversity have been drawn up as part of the REVERSE project, in which UIC has worked with its members and UKCEH to formulate a collective vision for protecting and enhancing biodiversity across the European rail network. They now form part of the European Railways: Strategy and Action Guide to ensure management for biodiversity is embedded at every level of the railway business, alongside safety, performance and sustainability. The adoption of the guidelines by member companies will be promoted through various UIC meetings and online events.
The REVERSE project comprises more than 20 European rail companies including Network Rail and SNCF as well as WWF (Worldwide Fund for Nature).
In 2021, UKCEH worked with Network Rail to draw up the rail company’s Biodiversity Action Plan to inform lineside habitat management across the UK. This involved using high-resolution imagery from satellites and aircraft to produce a detailed national map of all the habitats alongside the rail network.
At the moment Karin and I are in the UK for a couple of weeks. I had work to do as an external examiner at the University of Swansea, plus we wanted to catch up with some family and friends. Our main base has been the home of our mates Ian and Simone and we’ve enjoyed some warm, muggy evenings sitting in their garden chewing the fat. Every now and again my eyes have been drawn to the activities of bumblebees as they move in and out of the foliage of a small Silver Fir. The bees are attracted to the large colonies of an aphid that is feeding on the tree’s trunk, from which they are collecting honeydew, as you can see in the photograph above.
When we think of the diets of bees we automatically think “nectar and pollen”. Honeydew, as a sugar-rich fluid, fits broadly into this concept, though as far as I know there’s been little study of its relative importance as a food source for bees. Aside from a few “vulture bees”, all of the 20,000 or so species are vegetarian. And therein lies a problem. Bees evolved from carnivorous wasps and so the evolution of bees, and their complex ecologies, is tied into this profound dietary shift toward a plant-based diet.
A particular issue that has hardly been investigated until recently is that the ratio of elements within meat is very different to that of plants. In particular, animal tissue has a high ratio of sodium (Na) relative to potassium (K), whereas for plants the ratio is reversed – high ratio of K:Na.
In a new conceptual review paper with my colleagues Zuzanna Filipiak and Michał Filipiak, we have explored the implications of this difference in elemental ratios for bee ecology and evolution, and for the conservation of these important insects. The paper is open access and you can download a copy by following a link in this reference:
Bees provide important ecological services, and many species are threatened globally, yet our knowledge of wild bee ecology and evolution is limited. While evolving from carnivorous ancestors, bees had to develop strategies for coping with limitations imposed on them by a plant-based diet, with nectar providing energy and essential amino acids and pollen as an extraordinary, protein- and lipid-rich food nutritionally similar to animal tissues. Both nectar and pollen display one characteristic common to plants, a high ratio of potassium to sodium (K:Na), potentially leading to bee underdevelopment, health problems, and death. We discuss why and how the ratio of K:Na contributes to bee ecology and evolution and how considering this factor in future studies will provide new knowledge, more accurately depicting the relationship of bees with their environments. Such knowledge is essential for understanding how plants and bees function and interact and is needed to effectively protect wild bees.
As part of our roles as ambassadors of the new conservation organisation Restore (more of which later this year), several of us including Dave Goulson, George McGavin, and myself, are promoting this online petition to get the government to take the issue of neonicotinoid pesticides seriously. Here’s some text from Dave explaining the situation with a link to a petition that you can sign:
“For three years in a row our government has granted farmers special permission to use banned neonicotinoid pesticides on sugar beet. This is contrary to the expert advice of their own Expert Committee on Pesticides, who specifically recommended that permission should not be granted. It also flies in the face of a huge body of scientific evidence showing that these chemicals are phenomenally toxic to all insect life, and that their use on any crop contaminates soils, hedgerow plants, and nearby streams and ponds for years to come. We are in a crisis, with insect populations in freefall. It is about time our government woke up to this, and acted accordingly. This petition https://petition.parliament.uk/petitions/631948 is a necessary means of holding the government to account. Please sign and share, as signing will ensure the issue is debated in Parliament.”
This petition now has more than 15,000 signatures which ensures that it gets a response from the Government. If it reaches 100,000 mark, it will trigger a debate in Parliament. Please sign and promote this important initiative!
Note: This is a guest blog post by Guthrie Allen who is a PhD candidate at the University of East Anglia.
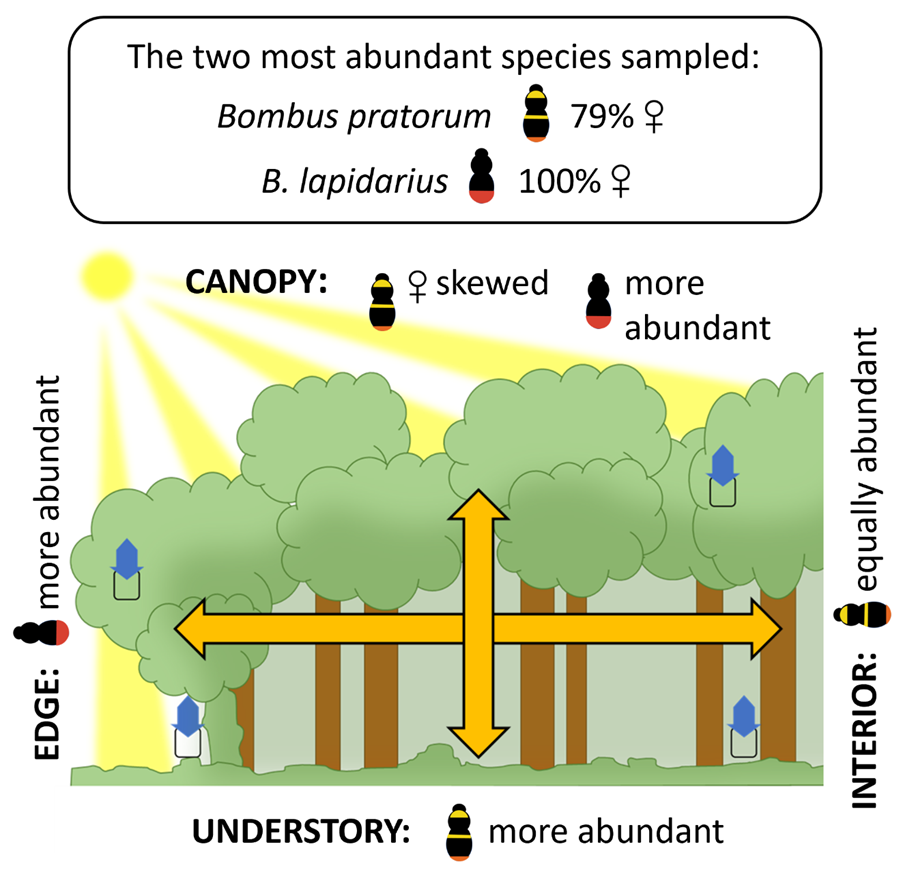
Though often overlooked, woodlands have great potential to support wild bees in farmed landscapes. Both the understory and canopy can provide large quantities of early-season nectar. The pollen of woodland-tree taxa such as Acer can form a large proportion of the early-season diets of bumblebees and solitary bees. Even the pollen from wind-pollinated trees such as Quercus can form a significant part of the diets of several common solitary bee species. Woodlands are also believed to provide ample nesting opportunities. These factors may explain why several studies have found positive correlations between bee abundance and/or diversity and woodland cover at the landscape scale. Despite this, a large-scale European study has shown that interior woodland understories support very few bees compared to exposed woodland edges.
Sampling the canopy – at heights of up to 20 metres – is not easy, but unlike the understory, this habitat remains sun-exposed after canopy closure and could be favourable for bees. In Europe, however, our knowledge of bee activity in the woodland canopy is very limited. Furthermore, whilst canopy-tree taxa have been identified as suitable food sources for many bees, we have little evidence that these trees are used for forage when located within woodlands. To explore the role of the canopy, we trapped bees in late spring in the understory and canopy – at the exposed edges and in the interiors – of 15 woodland sites across an English agricultural landscape.
A significant proportion of bee abundance was found in the canopy, represented by 23 of the 29 total sampled species. Interior canopy activity was much higher when nectar-producing Sycamore (Acer pseudoplatanus) trees were nearby. Communities differed between the canopy and understory: of the seven most common species sampled, three were more active in the understory, while the opposite was true of one species – Bombus lapidarius. Interestingly, we found the sex ratio of the most abundant species – B. pratorum – to be female-skewed in the canopy. For four of these species, and contrary to expectations, we found no evidence that understory activity was any higher at woodland edges compared to woodland interiors.
Further research is needed to understand community differences between the canopy and understory. Nonetheless, our results suggest that both these habitats have a significant role to play in supporting farmland bees. We demonstrate that a diverse bee community has the potential to exploit canopy resources, with nectar-producing trees in woodland interiors likely to provide forage for many bees. And we show the unexpected potential of interior woodland understories to support bee abundance.
Here’s the full reference with a link to the open access paper:
Allen, G. & Davies, R.G. (2022) Canopy sampling reveals hidden potential value of woodland trees for wild bee assemblages. Insect Conservation and Diversity. Available from: https://doi.org/10.1111/icad.12606
Here’s the abstract:
Woodlands can play an important role in supporting bee abundance and diversity in agricultural landscapes. However, in temperate-region studies, the canopy is rarely sampled, and our understanding of its contribution is limited. To explore this, we sampled bees in late spring with blue vane traps in the understory (n = 30) and crowns of mature Quercus robur (n = 35) at the exposed southern edges and in the interiors (ca. 25–75 m from woodland edges) of 15 woodland sites across an English agricultural landscape. A significant proportion of bee abundance and diversity was found in the canopy: canopy-trap catches were estimated to be a third as large as understory-trap catches, and 23 of the 29 sampled species were present in the canopy. Of the seven most common species sampled, four were equally abundant in woodland edge and interior traps; three were more abundant in understory traps, and a single species—Bombus lapidarius—was more abundant in canopy traps. The sex ratio of the most abundant species, B. pratorum, was female-skewed in the canopy. Additionally, the presence of nearby Acer pseudoplatanus trees in flower greatly increased canopy-trap catches in woodland interiors. These results suggest that both the woodland canopy and understory have a significant role to play in supporting farmland bee communities; they indicate the importance of nectar-producing trees in woodland interiors, opening avenues for canopy-based management; and they demonstrate that a diverse bee community has the potential to exploit canopy floral resources.
Most of us have at some time stared in fascination at the life contained within the pools that form on rocky shores at low tide. But none of us realized that a whole new class of ecological interaction was taking place!
The 12,000 or so described (and many un-named) seaweeds are incredibly important organisms. Their diverse and abundant photosynthesizing fronds make them one of the main primary producers in coastal seas, creating food and habitat for a huge range of animals. Not only that, but some – the coralline seaweeds – lock up vast amount of CO2 as calcium carbonate and help to create reef systems in the same way as coral.
Although scientists have studied seaweeds for hundreds of years, many aspects of their ecology are still unknown. Their detailed mode of reproduction, for example has only been studied in a small proportion of species.
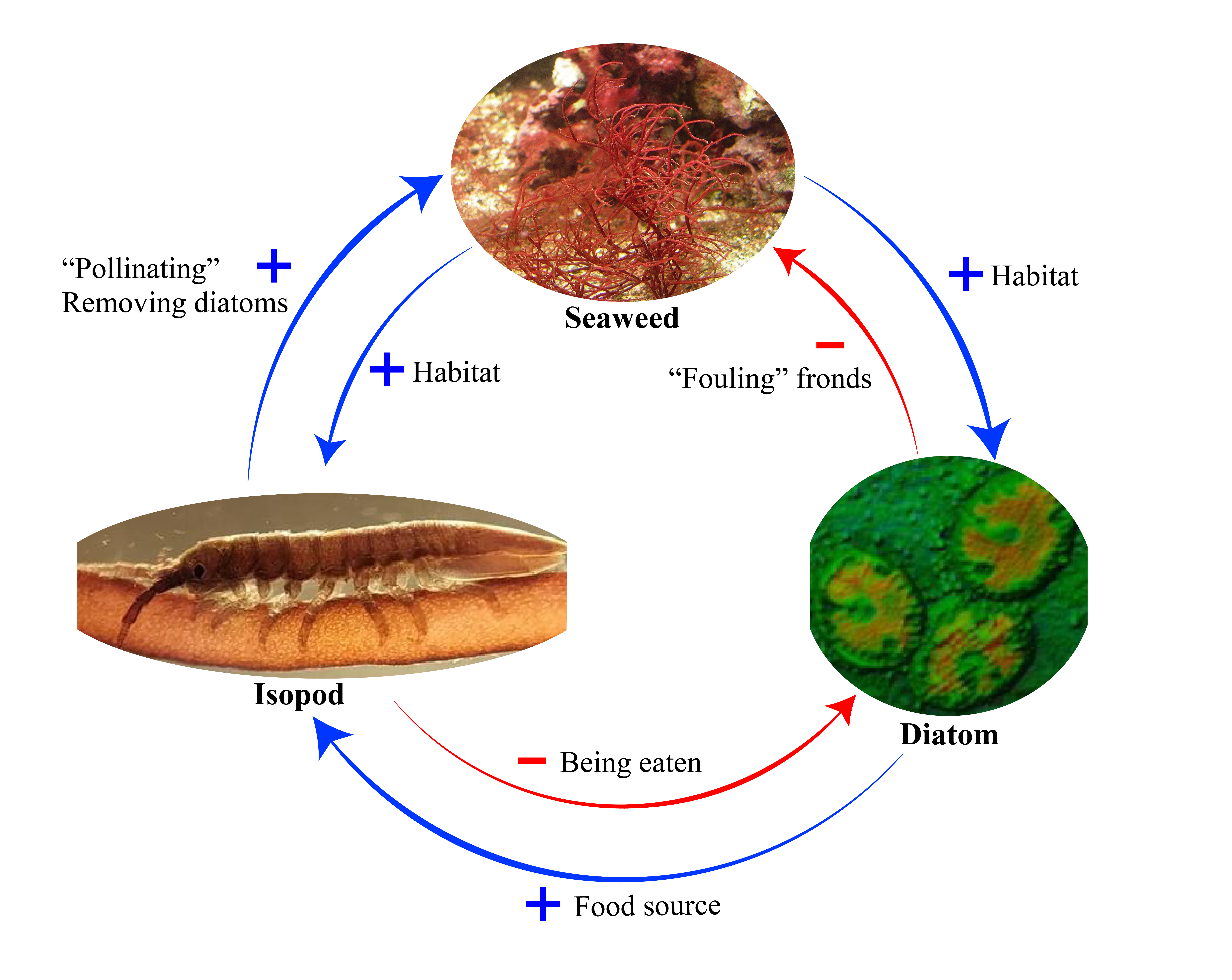
In a newly published study in the journal Science, French PhD researcher Emma Lavaut and her colleagues have shown that small isopod crustaceans – relatives of woodlice and sea slaters – facilitate the movement of the equivalent of seaweed sperm (termed “spermatia”) from male to female reproductive structures in just the same way that bees and other pollinators move pollen between flowers, so fertilizing female gametes.
Your read that correctly: some seaweeds have pollinators!
It’s an incredible finding! And the implications of this are enormous: Emma and her colleagues have added a whole new branch of life to the examples of sedentary (fixed-place) organisms that require a third party to enable their reproduction. In addition to being a fascinating biological discovery, it has significant environmental and sustainability implications.
Seaweeds are a diverse group of macroalgae that appeared more than one billion years ago, at least 500 million years before the evolution of what we think of as “true” plants, such as the flowering plants, conifers, cycads, ferns and mosses. Sexual reproduction in the brown and green seaweeds, which include kelps, wracks and sea lettuces, involves spermatia that are mobile and use a flagellum to swim through the water to seek out female reproductive structures. However, Emma studied a seaweed, Gracilaria gracilis, which belongs to the Rhodophyta or red seaweeds, and none of the species in this group have these swimming sperm equivalents.
Sexual reproduction in the red seaweeds has therefore always been something of a mystery. Three quarters of species have separate male and female individuals and so they cannot mate with themselves. It was assumed that the gametes were just released into water currents that haphazardly transported them to the female reproductive organs, much as wind pollinated grasses and pine trees release their vast clouds of pollen on land. The authors of this new study, however, point out that most sexual reproduction by these red seaweeds takes place in the relatively still waters of rock pools, a habitat that they mimicked in the laboratory in a series of elegant aquarium experiments.
The isopod crustaceans are attracted to the seaweed because they provide a habitat away from predators and a supply of food: they graze on the microalgae that colonise the seaweed’s fronds. Picking up spermatia and moving them between fronds is a side-effect of this activity by the small invertebrates. As you can see from the illustration above, the isopods and the seaweed are engaged in a “double mutualism“: a plus sign (+) indicates a positive effect of one species on another, while a minus sign (-) indicates a negative impact.
What I find especially fascinating about this research is that both the seaweed (Gracilaria gracilis) and the isopod (Idotea balthica) were originally described as species more than 200 years ago. They also have an extremely wide distribution. The isopod is found around the coasts of Europe and down the eastern seaboard of the Americas. The seaweed is pretty much found globally. These are not rare, unusual species, yet the interaction between them has only just been discovered! This is a point that I made in my recent book Pollinators & Pollination: Nature and Society: quite often, species that are well known interact in previously undocumented ways because no one has had the time or inspiration to look closely at them.
Although the idea that small sea creatures might be helping seaweeds to reproduce sounds very fanciful, there is a precedence for this discovery. Back in 2016, in a paper published in Nature Communications, a group of Mexican researchers led by Brigitta van Tussenbroek showed that a species of seagrass is pollinated by a diverse assemblage of small crustaceans and polychaete worms. Seagrasses are flowering plants, not seaweeds, but clearly this type of mutually beneficial relationship can exist between different species in the oceans.
Rhodophyta are the most diverse group of seaweeds, with more than 7,000 known species. They are especially abundant on coastal shores, oceanic habitats that are under huge pressure from infrastructure development, pollution, and climate change. At the same time, these seaweeds are economically important and millions of tonnes of them are collected every year as food, as nutritional and pharmaceutical supplements, and to produce agar. In order to conserve these seaweed populations, we need to better understand their ecology and their environmental requirements.
The work by Emma Lavaut and colleagues suggests that interactions with their “pollinators” may be a critical aspect of this understanding. In the same way that “Save the Bees” has been a rallying call for conserving interactions between species on land, we may soon hear this message echoed in “Save the Isopods”. At the very least, I have to add a new section to the second edition of my book!
Full disclosure: I was one of the reviewers of the original manuscript submitted to Science by Emma and her co-authors. It’s a rare privilege to review a study and think: “Wow! This is a game-changer!” and including this paper it’s happened to me only a handful of times. The editors at Science kindly invited my colleague Dr Zong-Xin Ren and myself to write a Perspective piece about the work and we were delighted to do so.
Image credits: Isopod and diatom images from Lavaut et al (2022). Gracilaria image by Emoody26 at English Wikipedia CC BY 3.0 https://commons.wikimedia.org/w/index.php?curid=3455016. Design by Shijia Wen and Jeff Ollerton.
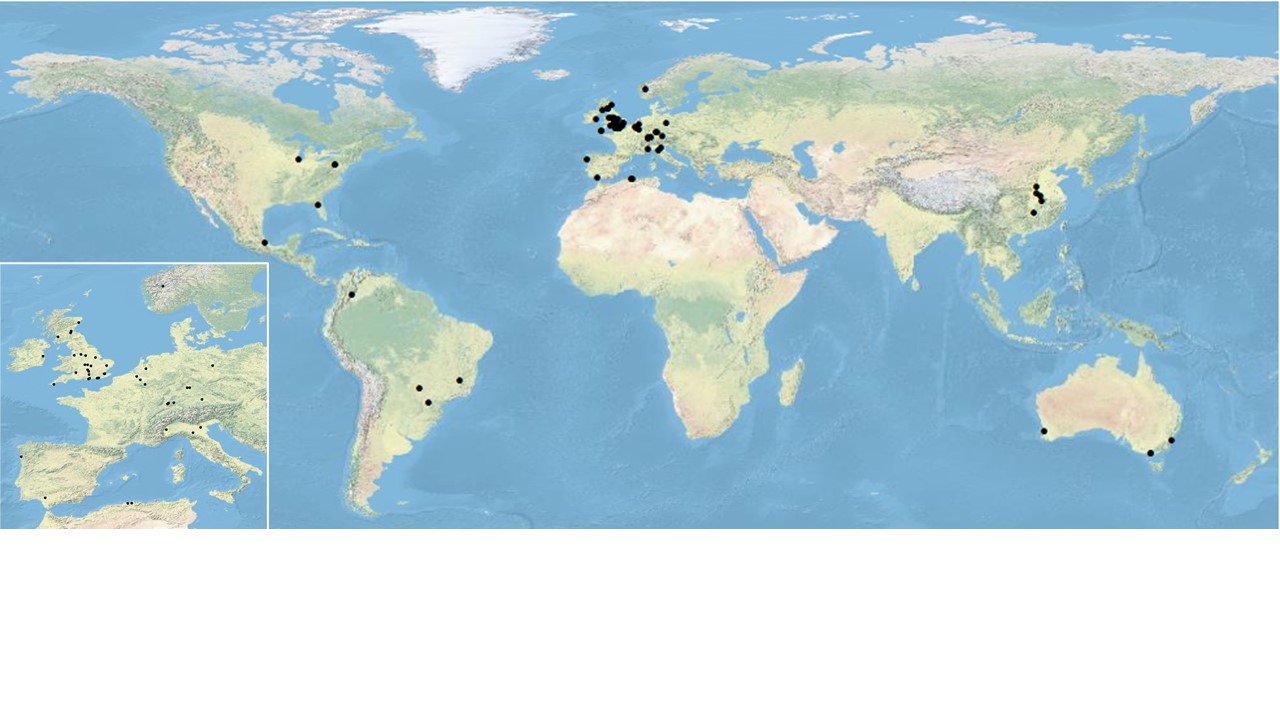
During the lockdown period of the COVID-19 pandemic in 2020, many pollination ecologists were stuck at home: universities and research institutes were closed and restrictions on travel meant that it was not possible to get out and do field work. In order to keep active and motivated, and to turn adversity into an opportunity, an ad hoc network of more than 70 researchers from 15 different countries (see the map above) decided to collect standardised data on the plant-pollinator networks in their own gardens and nearby public spaces.
When combined with information about location, size of garden, floral diversity, how the garden is managed, and so forth, this would provide some useful data about how gardens support pollinators. For those with kids at home it could also be a good way of getting them out into fresh air and giving them something to do!
The resulting data set of almost 47,000 visits by insects and birds to flowers, as well as information about flowers that were never visited, is freely available and will be an invaluable resource for pollination ecologists. For example, analysing the links between ornamental flowers that share pollinators with fruits and vegetables such as apples and beans, will allow us to make recommendations for the best plants to grow in home gardens that can increase yields of crops.
There’s an old saying about turning adversity into a positive outcome: “When life gives you lemons, make lemonade”, and the researchers were pleased to find that there’s one record of Citrus limon in the data set!
The paper describing the data set has just been published in the Journal of Pollination Ecology and you can download a PDF of the paper and the associated data for free by following this link.
Sincere thanks to all of my co-authors for their commitment to the project!
One of the projects with which I’ve been involved over the last year has been advising on a new book for children about bees and other pollinators, called Can We Really Help The Bees? Written by Katie Daynes and wonderfully illustrated by Róisín Hahessy, it tells the story of what happens when a swarm of bees comes to the window to let a group of children know that they, and their friends the other pollinators, are in trouble. Can they help? Yes they can!
It’s been a real pleasure working with Katie and Róisín on this project for Usborne Publishing and seeing the ideas, text, and illustrations evolve over time. I’ve written a short post over at the Usborne blog with some ideas about how to get children involved in helping the pollinators, and I think that it’s worth repeating one of the things that I wrote: everyone can make a difference to the wildlife around us and no one is too young to be involved!
Because of my involvement with Can We Really Help The Bees? I wasn’t able to include it on my curated list of the best books about bees and other pollinators at the Shepherd site. But it definitely should be on there and is highly recommended!
In my recent book Pollinators & Pollination: Nature and Society I discussed the current state of our knowledge of how populations of pollinators have changed over time. Although we have some quite detailed data for particular, often charismatic, species or for certain geographic localities or regions, for most species we know almost nothing. As I wrote in the chapter “The shifting fates of pollinators”:
“For most pollinators we are ‘data deficient’, in other words, we don’t know how their populations are performing. They could be doing well, but they may not be”
This is particularly true for those regions for the world that hold the greatest terrestrial biodiversity: the tropics. For the vast majority of species in the tropics we know precious little about trends in their populations and how their distributions have changed over time in the face of wide-scale land transformation and recent climatic shifts. Filling in some of the gaps in our knowledge of Neotropical pollinator distributions is one of its aims of SURPASS2, a collaboration between South American and UK ecologists, and one of several research and outreach projects with which I’m involved.
In a new study that’s come out of that work, led by Rob Boyd from the UK Centre for Ecology and Hydrology, we’ve used the GBIF database to look at the changing distributions of four important groups of pollinators: bees, hoverflies, leaf-nosed bats and hummingbirds. In particular we were interested in understanding the kinds of biases that come with such publicly available data, and whether recent efforts to add data to GBIF has improved our understanding of trends.
Our overall conclusion is that there are significant limitations and biases inherent in all of these data sets even for groups like hummingbirds which one would imagine are well documented by scientists and bird-watching naturalists. In addition, having more data does not necessarily help matters: it can introduce its own biases.
The paper is open access and feely available; here’s the reference with a link:
Aim Aggregated species occurrence data are increasingly accessible through public databases for the analysis of temporal trends in the geographic distributions of species. However, biases in these data present challenges for statistical inference. We assessed potential biases in data available through GBIF on the occurrences of four flower-visiting taxa: bees (Anthophila), hoverflies (Syrphidae), leaf-nosed bats (Phyllostomidae) and hummingbirds (Trochilidae). We also assessed whether and to what extent data mobilization efforts improved our ability to estimate trends in species’ distributions.
Location The Neotropics.
Methods We used five data-driven heuristics to screen the data for potential geographic, temporal and taxonomic biases. We began with a continental-scale assessment of the data for all four taxa. We then identified two recent data mobilization efforts (2021) that drastically increased the quantity of records of bees collected in Chile available through GBIF. We compared the dataset before and after the addition of these new records in terms of their biases and estimated trends in species’ distributions.
Results We found evidence of potential sampling biases for all taxa. The addition of newly-mobilized records of bees in Chile decreased some biases but introduced others. Despite increasing the quantity of data for bees in Chile sixfold, estimates of trends in species’ distributions derived using the postmobilization dataset were broadly similar to what would have been estimated before their introduction, albeit more precise.
Main conclusions Our results highlight the challenges associated with drawing robust inferences about trends in species’ distributions using publicly available data. Mobilizing historic records will not always enable trend estimation because more data do not necessarily equal less bias. Analysts should carefully assess their data before conducting analyses: this might enable the estimation of more robust trends and help to identify strategies for effective data mobilization. Our study also reinforces the need for targeted monitoring of pollinators worldwide.
——————————————————–
SURPASS2 has been a hugely productive project as you’ll see if you look at the Publications page of the website. There’s much more to come and I’ll report on those research papers as they appear.
First the misleading title. This ‘debunks’ claim actually compares two different things: 75% of CROPS being dependent on pollinators versus 10% of crop YIELD. However, even if we focus on the 10% claim, a small increase in yield can be the difference between profit and bankruptcy for small-scale farmers. And most of the world’s farmers are small-scale and living on the borderline between loss and break-even. In addition, there’s no acknowledgement of the food production from home gardens, allotments, and community gardens, which is significant but largely unquantified.
Next, by focusing on yield and comparing, say, wind-pollinated wheat with insect-pollinated apples, the article takes no account of the fact that many of these crops that depend to some extent on pollinators mainly provide essential vitamins and minerals – not calories – to diets. When I tweeted about this earlier in the week, one person commented that they describe the insect-pollinated foods as ‘an important source of flavour and colour in our diets, rice and wheat are all well and good, but you do kinda need something more than grey slop to live’. Another said: ‘I’m so glad you mentioned this. I’m sick of reading articles that praise innovations to increase calories, when what we need is better nutrition from vitamins, minerals & fibres’.
Both great points, and well made.
That essay was also factually incorrect when it described roots crops such as carrots or some of the leafy cabbages and lettuces as not requiring pollinators. Many varieties of these crops ARE pollinator dependent: how do they think we get the seed for the next year’s crop?! And there are many crops and varieties that have not been evaluated for their dependency on pollinators: the 75% figure actually refers to the 115 most productive crop plants (Klein et al. 2007).
When I tweeted about the essay I commented that I was very disappointed by ‘Our World in Data’ – they are usually better than this when it comes to the facts. What I hadn’t appreciated at the time was that in fact the Genetic Literacy Project had highjacked the original piece by Hannah Ritchie and reworked it to give it a very different slant*.
This is where it starts to get dishonest and in fact the Genetic Literacy Project (GLP) has form in this area. The Sourcewatch site describes the GLP as ‘a corporate front group that was formerly funded by Monsanto’ with a remit to ‘shame scientists and highlight information helpful to Monsanto and other chemical producers’. In other words it’s heavily tied to Big Agriculture which, of course, would like us to believe that there’s not an issue with declining pollinators, that pesticides and agricultural intensification are our friends, and that Everything Is OK. Read the full account here.
Frankly, the GLP is so tainted that I’d not believe anything that they publish.
Pollinator decline and the role of pollinators in agriculture are complex issues. If you’d like to know more about the importance of pollinators to agriculture, complete with some accurate and objective facts, then there’s a whole chapter devoted to the topic in my book Pollinators & Pollination: Nature and Society.
*Note that I’ve been communicating with Hannah about the root and leaf crop issue and she accepts that this needs to change in the original. She’s also asked the Genetic Literacy Project to take down their version as it contravenes copyright.
Reference
Klein, A.-M., Vaissière, B.E., Cane, J.H. et al. (2007) Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society of London B 274: 303–313.