Although I’m one of the speakers, I can’t make it in person as I’ve got teaching commitments at Roskilde University, so I’ve recorded my ten minute piece and I’m happy to answer questions via the Contact page on my website.
It’s been a couple of years since I last did a talk or workshop for the Wildlife Trust for Bedfordshire, Cambridgeshire & Northamptonshire. But I’m pleased to say that they’ve invited me back and you can join me tomorrow evening for an online introductory talk about pollinators and pollination in the UK.
The talk starts at 7pm UK time and full details of how to sign up are in the link below:
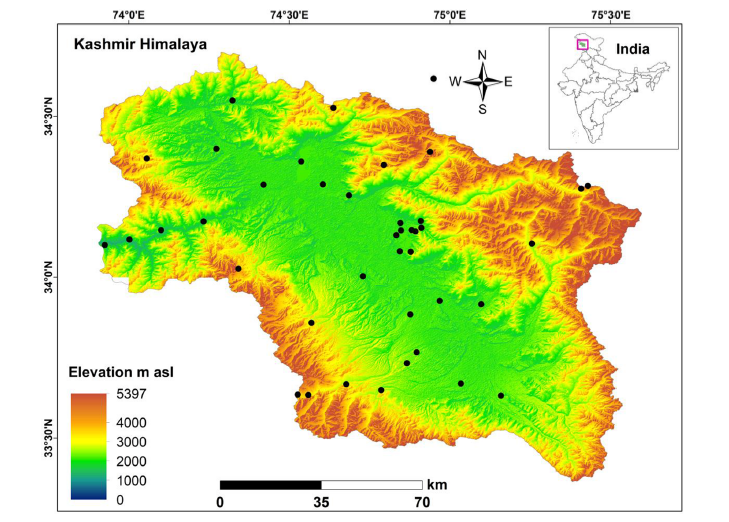
Although we sometimes like to think that we have a “global” perspective on plant-pollinator interactions, in truth there are large parts of the world where we have little or no information. That’s especially true of biodiversity hotspots. One such hotspot is the Kashmir Himalaya, a relatively small area (just 135 km long by 32 km wide) that nonetheless contains an estimated 2,000 species of flowering plant, more than 150 of which are endemic to the area. In addition, over 1,280 insect pollinator species have been described, including at least 29 species of bumblebees and as many as 40 species of butterflies.
Recently I’ve been collaborating with Dr Zubair Rather from the University of Kashmir and his colleagues on a data set that represents the first network analysis of plant-pollinator interactions from the region. The network is presented at a large spatial scale – what is often termed a “meta-network”. As Dr Kit Prendergast and I noted in our recent paper, scale matters when it comes to analysing these sorts of networks. Nonetheless (to quote the current paper):
“meta-networks represent the “backbone” of plant-pollinator interactions to which smaller, local networks are attached”
Even at this very large scale we’re seeing some fascinating patterns emerging with respect to the plants, for example the importance of cultivated apples in supporting the wider network of flower visitors. Also notable, and I believe demonstrated for the first time, is the fact that both the indigenous Asian Honeybee (Apis cerana) and the introduced Western Honeybee (A. mellifera) utilise exactly the same, extremely broad array of plants from which to collect nectar and pollen.
Our Kashmir meta-network is, hopefully, the starting point for further work on plant-pollinator interactions in this fascinating and diverse part of the world. The paper is published in a special issue of the journal Flora that’s dedicated to the importance of natural history when considering the ecology and evolution of plant-pollinator relationships. The special issue is a celebration of the work of Professor Marlies Sazima and is edited by Dr Pedro Bergamo.
Here’s the reference with a link to download a free copy which should be valid up to and including 23rd January 2023:
Plant-pollinator studies are increasingly using network analysis to investigate the structure and function of such communities. However, many areas of high biodiversity largely remain unexplored in this way. Our study describes a plant-pollinator meta-network from an understudied biodiversity hotspot, the Kashmir Himalaya, where we specifically investigate plant-pollinator network nestedness and modularity, as well as the influence of alien species and the impacts of simulating species extinctions on network structure. Natural history observations were used to document the meta-network between 230 plant and 80 pollinator species forming 1958 (11% of the possible) interactions. Among the plants Malus domestica and among the pollinators Apis mellifera and A. cerana formed the largest number of interactions with significant influence over the whole network. Network cumulative degree distribution depicted a higher number of degree levels in pollinators than plants. A moderately high number of realized interactions were revealed, thereby indicating potential structural and functional stability in the network. Eight strongly defined modules were observed in the network which varied in their composition. For example, the Ephedra module exclusively comprised of native species whereas the Apis module comprised of all the four different types of interacting species (i.e. native and alien plants and pollinators) and also integrated the highest number of alien species. In the network overall, 40% of interactions were by alien species, reflecting how well these were integrated. Extinction simulations suggested that the network would collapse more quickly when the most connected pollinators are removed, rather than the most connected plant species. Our study is the first assessment of a plant-pollinator network from this Himalayan biodiversity hotspot; and will help to inform the ecological and economic implications of plant-pollinator interactions in an era of global biodiversity crisis.
Note: This is a guest blog post by Guthrie Allen who is a PhD candidate at the University of East Anglia.
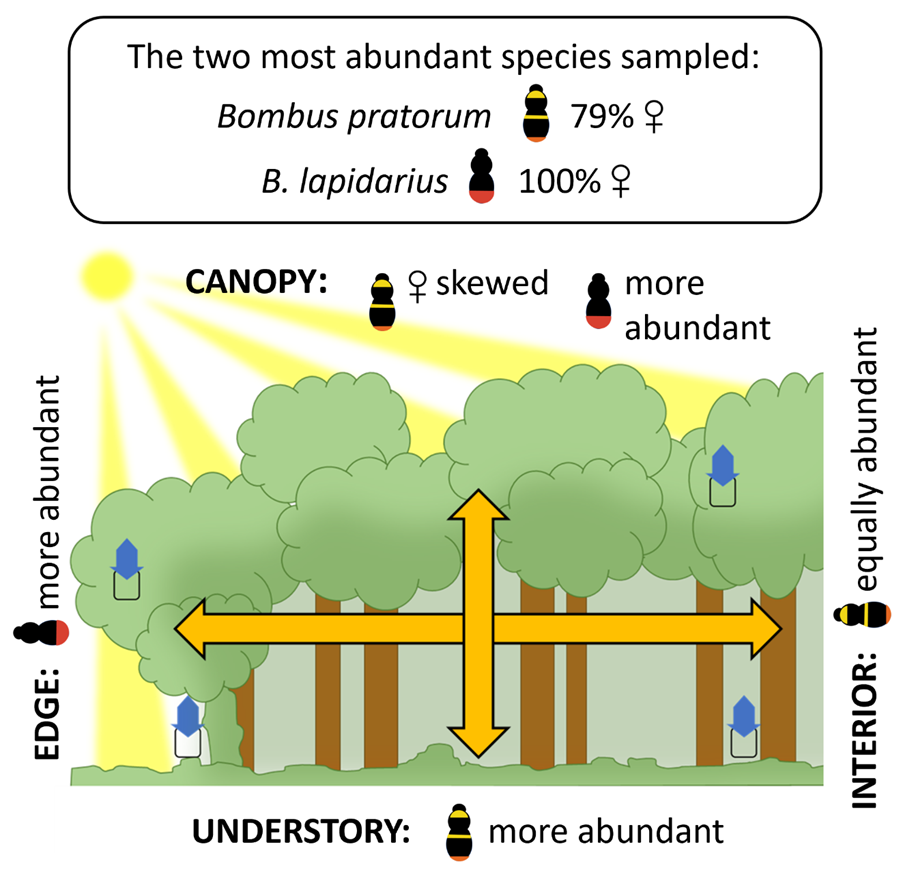
Though often overlooked, woodlands have great potential to support wild bees in farmed landscapes. Both the understory and canopy can provide large quantities of early-season nectar. The pollen of woodland-tree taxa such as Acer can form a large proportion of the early-season diets of bumblebees and solitary bees. Even the pollen from wind-pollinated trees such as Quercus can form a significant part of the diets of several common solitary bee species. Woodlands are also believed to provide ample nesting opportunities. These factors may explain why several studies have found positive correlations between bee abundance and/or diversity and woodland cover at the landscape scale. Despite this, a large-scale European study has shown that interior woodland understories support very few bees compared to exposed woodland edges.
Sampling the canopy – at heights of up to 20 metres – is not easy, but unlike the understory, this habitat remains sun-exposed after canopy closure and could be favourable for bees. In Europe, however, our knowledge of bee activity in the woodland canopy is very limited. Furthermore, whilst canopy-tree taxa have been identified as suitable food sources for many bees, we have little evidence that these trees are used for forage when located within woodlands. To explore the role of the canopy, we trapped bees in late spring in the understory and canopy – at the exposed edges and in the interiors – of 15 woodland sites across an English agricultural landscape.
A significant proportion of bee abundance was found in the canopy, represented by 23 of the 29 total sampled species. Interior canopy activity was much higher when nectar-producing Sycamore (Acer pseudoplatanus) trees were nearby. Communities differed between the canopy and understory: of the seven most common species sampled, three were more active in the understory, while the opposite was true of one species – Bombus lapidarius. Interestingly, we found the sex ratio of the most abundant species – B. pratorum – to be female-skewed in the canopy. For four of these species, and contrary to expectations, we found no evidence that understory activity was any higher at woodland edges compared to woodland interiors.
Further research is needed to understand community differences between the canopy and understory. Nonetheless, our results suggest that both these habitats have a significant role to play in supporting farmland bees. We demonstrate that a diverse bee community has the potential to exploit canopy resources, with nectar-producing trees in woodland interiors likely to provide forage for many bees. And we show the unexpected potential of interior woodland understories to support bee abundance.
Here’s the full reference with a link to the open access paper:
Allen, G. & Davies, R.G. (2022) Canopy sampling reveals hidden potential value of woodland trees for wild bee assemblages. Insect Conservation and Diversity. Available from: https://doi.org/10.1111/icad.12606
Here’s the abstract:
Woodlands can play an important role in supporting bee abundance and diversity in agricultural landscapes. However, in temperate-region studies, the canopy is rarely sampled, and our understanding of its contribution is limited. To explore this, we sampled bees in late spring with blue vane traps in the understory (n = 30) and crowns of mature Quercus robur (n = 35) at the exposed southern edges and in the interiors (ca. 25–75 m from woodland edges) of 15 woodland sites across an English agricultural landscape. A significant proportion of bee abundance and diversity was found in the canopy: canopy-trap catches were estimated to be a third as large as understory-trap catches, and 23 of the 29 sampled species were present in the canopy. Of the seven most common species sampled, four were equally abundant in woodland edge and interior traps; three were more abundant in understory traps, and a single species—Bombus lapidarius—was more abundant in canopy traps. The sex ratio of the most abundant species, B. pratorum, was female-skewed in the canopy. Additionally, the presence of nearby Acer pseudoplatanus trees in flower greatly increased canopy-trap catches in woodland interiors. These results suggest that both the woodland canopy and understory have a significant role to play in supporting farmland bee communities; they indicate the importance of nectar-producing trees in woodland interiors, opening avenues for canopy-based management; and they demonstrate that a diverse bee community has the potential to exploit canopy floral resources.
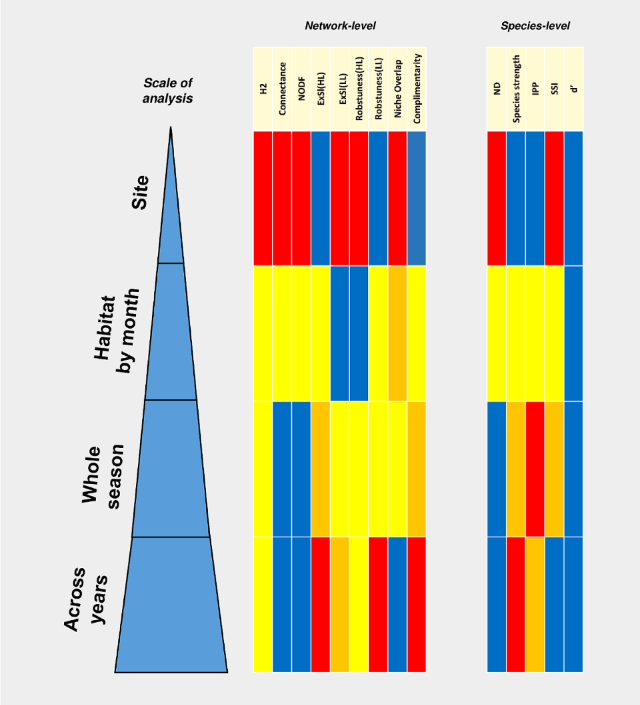
It’s long been recognised that the scale at which we study the natural world – over long or short time periods, or across small areas or whole regions – affects the conclusions that we draw about ecological patterns and processes. This is certainly true of plant-pollinator interactions. For example, a widely distributed plant can have very different pollinators at the extremes of its range, and pollinators like bees may vary their focus on nectar and pollen sources from year to year.
The analysis of these interactions as networks of actors has become increasingly popular in the last couple of decades. However there is no consensus about how frequent sampling should be, or the geographical scale over which networks should be studied. In fact all scales (from regional “meta-networks” down to single-season, single-site, single taxon observations) are relevant, depending on the questions being asked or the hypotheses posed.
But it’s important that we acknowledge that conclusions drawn at one scale may not apply at other scales.
That’s the take home message from a paper published this week which is the latest output from the PhD work of Australian bee expert Kit Prendergast. We have collaborated on several papers based on her data and this is actually my 100th peer-reviewed publication: a proud milestone for me and one which I’m glad to share with a wonderful early career researcher like Kit!
Here’s the reference with a link to a read-only version of the paper:
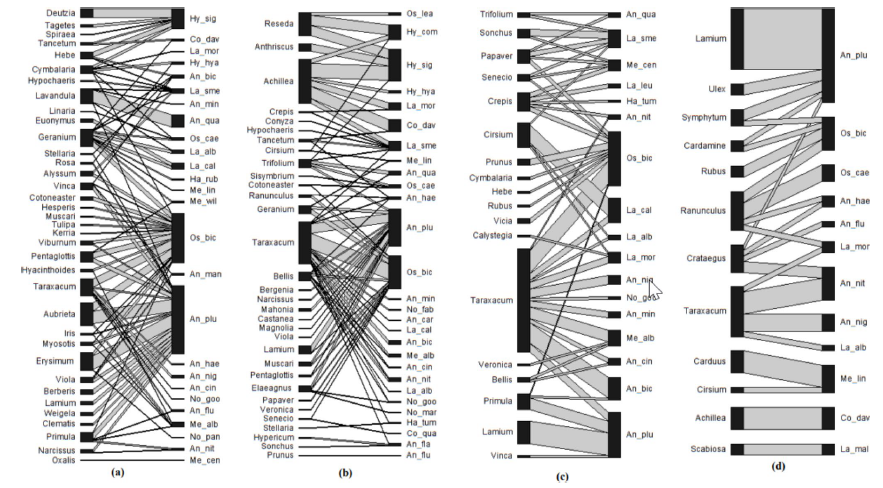
Bipartite networks of flowering plants and their visitors (potential pollinators) are increasingly being used in studies of the structure and function of these ecological interactions. Whilst they hold much promise in understanding the ecology of plant– pollinator networks and how this may be altered by environmental perturbations, like land-use change and invasive species, there is no consensus about the scale at which such networks should be constructed and analysed. Ecologists, however, have emphasised that many processes are scale dependent. Here, we compare network- and species-level properties of ecological networks analysed at the level of a site, pooling across sites within a given habitat for each month of surveys, and pooling across all sites and months to create a single network per habitat type. We additionally considered how these three scales of resolution influenced conclusions regarding differences between networks according to two contrasting habitat types (urban bushland remnants and residential gardens) and the influence of honey bee abundance on network properties. We found that most network properties varied markedly depending on the scale of analysis, as did the significance, or lack thereof, of habitat type and honey bee abundance on network properties. We caution against pooling across sites and months as this can create unrealistic links, invalidating conclusions on network structure. In conclusion, consideration of scale of analysis is also important when conducting and interpreting plant–pollinator networks.
Within the last decade there’s been a growing awareness of the importance of urban environments for supporting populations of pollinators, especially bees. Indeed, I devoted a whole chapter of my book Pollinators & Pollination: Nature and Society to the topic, though even then I was only able to scratch the surface of the research that’s been done. Since then there’s been some important studies published and this 2020 review by Kath Baldock provides a good starting point for the topic, whilst a recent pre-print by Pietro Maruyama and colleagues emphasises how little we know about pollinators in tropical cities.
One of the most detailed studies of urban solitary bees in a British town was conducted by Muzafar Sirohi when he was a PhD researcher in my department in Northampton. The first paper from that work, documenting the diversity and abundance of bees, came out in 2015, but since then commitments to other projects, plus Muzafar’s return to his university in Pakistan, have meant that we’ve struggled to find the time to publish more. Hopefully that’s changing and the second publication from Muzafar’s thesis is now out, with a third in progress.
This new paper uses a network approach to study the use of flowers by these bees; here’s the reference with a link to a read-only copy of the paper, followed by the abstract.
Biodiversity is declining through human activities and urbanisation is often seen as a particular concern. Urban settings, however, provide diverse microclimatic conditions for plants and pollinating insects, and therefore may be significant habitats for the conservation of solitary and primitively eusocial bees, a major group of pollinators. This study analysed the interactions between these bees and the plants on which they forage, using a network approach. We compared urban habitats (gardens, roadsides, and open vegetation) in a large British town with nearby nature reserves. One native plant Taraxacum officinale (dandelion) was a core generalist species visited in all habitat types. Other core plant species restricted to particular habitats include species of Geranium, Bellis, Crepis, and Ranunculus. Two generalist bee species, Anthophora plumipes and Osmia bicornis were the core visitor species within the networks. The networks were comparatively more nested in urban habitat types than nature areas, suggesting more frequent interactions between generalist and specialist species in urban areas. Network connectance, network level specialisation (H2’ index), and plant generality (network level) were not significantly different in urban and nature areas. However, visitor generality was found to be significantly higher in urban gardens than in nature areas. Careful management of common urban vegetation would be beneficial for supporting urban wild pollinators.
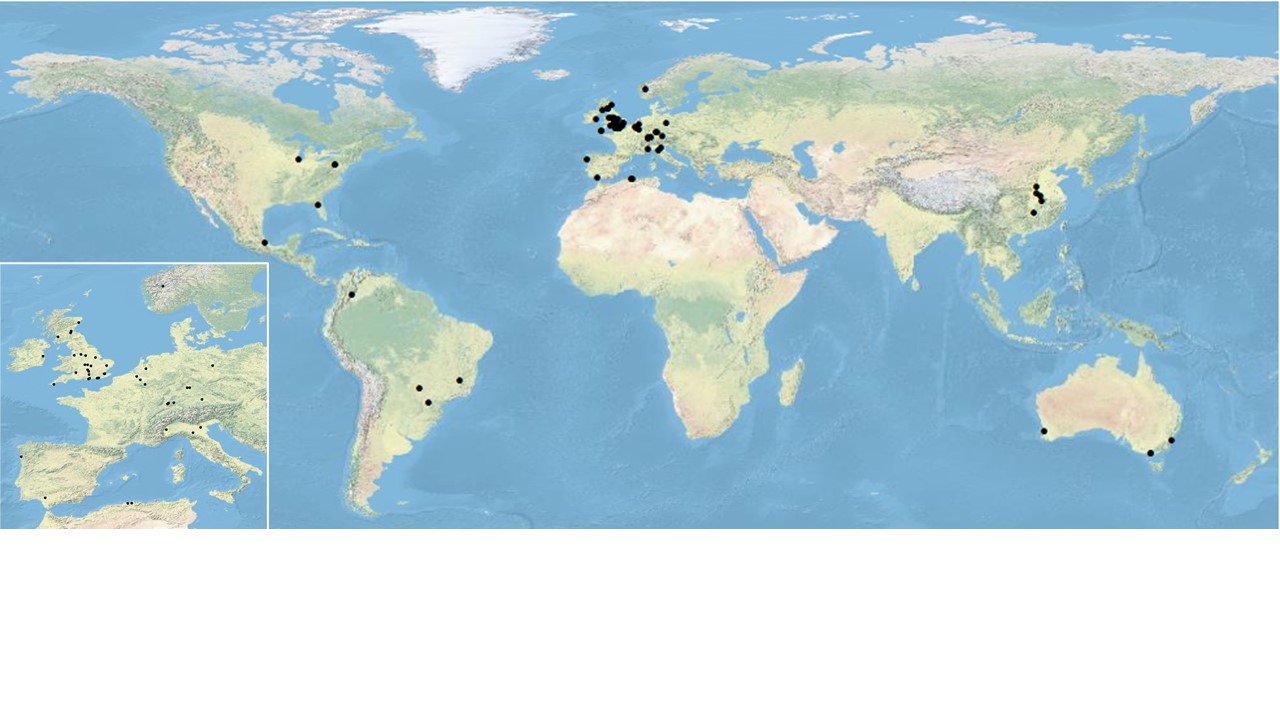
During the lockdown period of the COVID-19 pandemic in 2020, many pollination ecologists were stuck at home: universities and research institutes were closed and restrictions on travel meant that it was not possible to get out and do field work. In order to keep active and motivated, and to turn adversity into an opportunity, an ad hoc network of more than 70 researchers from 15 different countries (see the map above) decided to collect standardised data on the plant-pollinator networks in their own gardens and nearby public spaces.
When combined with information about location, size of garden, floral diversity, how the garden is managed, and so forth, this would provide some useful data about how gardens support pollinators. For those with kids at home it could also be a good way of getting them out into fresh air and giving them something to do!
The resulting data set of almost 47,000 visits by insects and birds to flowers, as well as information about flowers that were never visited, is freely available and will be an invaluable resource for pollination ecologists. For example, analysing the links between ornamental flowers that share pollinators with fruits and vegetables such as apples and beans, will allow us to make recommendations for the best plants to grow in home gardens that can increase yields of crops.
There’s an old saying about turning adversity into a positive outcome: “When life gives you lemons, make lemonade”, and the researchers were pleased to find that there’s one record of Citrus limon in the data set!
The paper describing the data set has just been published in the Journal of Pollination Ecology and you can download a PDF of the paper and the associated data for free by following this link.
Sincere thanks to all of my co-authors for their commitment to the project!
Over the weekend there was a discussion on Twitter about “beewashing” that was spun out of this tweet by London beekeeper Richard Glassborow. Richard and his colleagues are some of the most responsible beekeepers that I know and they are getting increasingly frustrated by claims from irresponsible companies that keeping a hive of bees in your garden will help to “save the bees”, backed up by spurious claims that “honeybee colonies are dying out”.
The Twitter exchange prompted me to produce the Condescending Wonka meme that you see above because, as I discussed in my recent book Pollinators & Pollination: Nature and Society, pollinator conservation is a really complex area. But there’s no doubt that beekeeping as it’s being widely promoted is not the answer to bee conservation. Let me explain why.
The word “honeybee” does not refer to just one species. It’s most often* applied to bees in the genus Apis, especially the Western Honeybee Apis mellifera, but there are another seven or so Apis species to which the word can be applied. Of those other Apis species, most have never been domesticated and they live as free-living colonies is the various parts of Asia where they evolved. Only Apis cerana is kept in hives, as far as I am aware. The conservation status of most of these other Apis species is unclear but given that they are predominantly forest species, and deforestation is a chronic problem in Asia, we can surmise that some species may be declining. If you want to know more about them the Wikipedia page is a good starting point.
In this short post I just want to consider the Western Honeybee (Apis mellifera). This is a really knotty species to get to grips with because there are multiple subspecies and within subspecies there are various genetic lineages. In addition, the Western Honeybee has been subject to artificial selection for desirable qualities, such as docility, amount of honey produced per hive, and disease resistance, as well as cross-breeding between different subspecies**. The best recent summary of our current understanding of Western Honeybee genetics and conservation is this 2019 review by Fabrice Requier and colleagues, from which I’ve drawn quite a bit of information.
For the purposes of this explaining what’s going on, it’s easiest to think about the species as comprising three “megapopulations”:
Western Honeybees that are managed in hives: For the most part these are not endangered. Britain has as many hives now as it did in the mid-1950s and indeed globally we have more hives than ever (about 90 million hives at the last count). They are found far beyond their natural range and have been introduced into places where they are not native such as the Americas, parts of Asia, and Australia. STATUS: doing just fine.
Western Honeybees that have founded “feral” colonies: These have escaped from hives in countries where they have been introduced and become naturalised. They are doing well, too well in fact: they are a significant conservation issue in places like Australia. STATUS: doing just fine.
Western Honeybees that are living wild in their native range: This is where things become a little muddier. The African populations of the various subspecies seem to be doing well, but more studies are needed to confirm this. In Europe, actually defining what constitutes “wild” honeybees across a region where a lot of selection and hybridization has gone on, probably for thousands of years, is tricky. However there’s no doubt that wild colonies of Apis mellifera are not uncommon in suitable woodland: see this paper about free-living colonies in Ireland by Keith Browne and colleagues, for instance. Note their statement that genetic evidence shows that “the free-living population sampled is largely comprised of pure A. m. mellifera“, i.e. the European Black Honeybee. STATUS: probably doing quite well though more data is needed.
Conclusion: as I said, it’s really complicated and I don’t pretend to have all of the answers, no one does. But what IS clear is that managed Western Honeybees are not declining and keeping yet more hives of them is not going to help us to “Save the Bees”. I’ll leave the last word to Requier et al., whose review I really do recommend: “We argue for the redirection of attention from managed honey bees to the neglected conservation of wild honey bees.” Amen to that.
———————————————————————————————
*The term “honeybee” is sometimes also used for other social bees that produce honey, for example stingless honeybees in the genus Trigona, but there’s no real consensus on what “honey” actually is, and as I’ve argued in another post, bumblebees (Bombus spp.) also produce honey.
As kids, my friends and I did a lot of digging. We always seemed to be burrowing into slopes or excavating trenches, pretending to be archaeologists or treasure hunters. Indeed, there was a lot of ground treasure to be found in the part of Sunderland where I grew up. The area has a long history of pottery and glass making, and ship building, and the remnants of these industries could be uncovered every time we stuck a spade in the earth. Over time I developed my own small museum of interesting, unearthed fragments, including bits of hand-painted ceramics, glass bottles, and unidentifiable metal shards, alongside various animal bones I’d excavated. My parents quietly indulged this interest, and my muck-streaked face and clothes, even if they didn’t quite understand what I was doing.
Aged about 10, my first encounter with a bumblebee nest was during one such dig. On the waste ground behind a large advertising hoarding, we began digging into a low, grass-covered mound and accidentally excavated what was probably a small nest of Buff-tailed Bumblebees (Bombus terrestris). I can recall being fascinated by the waxy, odd shaped cells and by the sticky fluid that some of them were leaking. Being an adventurous sort of child I tasted the liquid: it was sweet and sticky, and that was my first encounter with bumblebee “honey”.
I’m going to leave those quotation marks in place because if you do an online search for “do bumblebees make honey?” you generally find that the answer is “no, only honey bees make honey”.
Now, defining honey as something made by honey bee strikes me as a circular argument at best. And it also neglects the “honey” made by meliponine bees that is central to the culture of stingless bee keeping by indigenous groups in Central and South America, and the long tradition pre-colonial tradition of honey hunting by Aboriginal Australians. So if we widen our definition of “honey” as being the nectar*-derived fluid stored in the nests of social bees, then Apis honey bees, stingless bees and bumblebees must all, by logic, make honey. And likewise there’s wasps in the genus Brachygastra from Central and South America that are referred to as “honey wasps” because, well, I’m sure you can work it out!
But this is where things become a little trickier, because turning nectar* into honey involves some complex evaporation and enzymatic activity, so that the resulting fluid is more concentrated and dominated by the sugars glucose and fructose. Although analysis of honey bee honey is commonplace, and there’s been some research conducted on the honey of stingless bees, I don’t know of any studies that have compared Bombus honey with that of other bees, or with what is stored in the nests of honey wasps**. If I’ve missed anything, please do comment and let me know, but this strikes me as an area of research demanding some attention.
So do bumblebees make honey? That very much depends on our definitions, but I’m happy to accept that they do because “honey” is not a single thing: it’s an insect-derived substance that can take a range of forms but serves the same broad purpose of feeding the colony. And although insects have probably been producing it for millions of years, I think I’ve known the answer to the question for almost 50 of them…
UPDATE: A couple of people have commented on social media that there are legal definitions of “honey” as a foodstuff. Here’s the definition according to UK law***:
“the natural sweet substance produced by Apis mellifera bees from the nectar of plants or from secretions of living parts of plants or excretions of plant-sucking insects on the living parts of plants which the bees collect, transform by combining with specific substances of their own, deposit, dehydrate, store and leave in honeycombs to ripen and mature”
So, legally, we can’t call anything that isn’t made by Apis mellifera “honey”, at least from a foodstuffs regulation perspective. But that’s clearly different to what we have been discussing above, which is about a biological definition of honey.
It’s also interesting to look at the compositional requirements of honey as a foodstuff (presented in Schedule one of that document, if you follow the link above). The lower limit for moisture content is 20%. Now if you consider that most nectar in flowers has a sugar content of between about 20% and 50%, clearly there’s been a lot of evaporative work done by the bees to reduce the amount of water in the honey. I would love to know how bumblebee (and other insect) “honey” compares to this: do they put the same kind of effort into evaporating the water from the stored nectar? Given that the purpose of reducing the water content is to prevent fermentation by yeasts when it’s stored for a long time, and that there are bumblebee species which have colonies that are active for more than one year, I imagine that at least some species in some parts of their range may employ similar tactics.
Thanks to everyone who has been commenting and discussing the topic. It never ceases to amaze me how much we still do not understand about some fundamental aspects of the natural history of familiar species!
*And honeydew to a greater or lesser extent.
**I’m going to ignore honey pot ants for now as this is complex enough as it is and they don’t store the “honey” in nest cells.
***From what I can gather definitions in other countries are similar.
Although it can be invasive and an environmental nuisance in parts of the world where it’s introduced, common or European ivy (Hedera helix) is clearly one of the most vital plants across its native range of Europe, southern Scandinavia and the Mediterranean. Its clinging stems bind the landscape and provide habitat for a diversity of creatures. By offering nectar at a time when there’s few other plants in flower, and berries at a crucial point in the winter, ivy bridges a food gap for both nectar feeding insect and fruit eating birds and mammals.
Ivy is a very popular subject for student research because it’s in flower at the start of the university academic year. In the past I’ve had several students carry out their final year projects using ivy to test ideas about pollinator effectiveness and plant reproductive success. Because the open, densely-clustered flowers can dust pollen onto any insect that visits, the most effective pollinators will vary depending on which are abundant at any time and place, and include various types of flies and bees, plus those much-misunderstood wasps!
Perhaps we should leave the final word on ivy to the Northamptonshire ‘Peasant Poet’ John Clare who wrote ‘To the Ivy’ in the early 19th century:
Jacobs, J.H., Clark, S.J., Denholm, I., Goulson D., Stoate, C. & Osborne J.L. (2010) Pollinator effectiveness and fruit set in common ivy, Hedera helix (Araliaceae). Arthropod-Plant Interactions 4: 19–28
Ollerton, J. (2021) Pollinators & Pollination: Nature and Society. Pelagic Publishing, Exeter, UK
Ollerton, J., Killick, A., Lamborn, E., Watts, S. & Whiston, M. (2007) Multiple meanings and modes: on the many ways to be a generalist flower. Taxon 56: 717-728