But just as when a movie director says “That’s a wrap” at the end of the final day of filming, the hard work does not stop here. Two people have read the full manuscript as I was producing chapters and their suggestions have been incorporated into this draft. The publisher will now send it to a third, independent beta reader and once their feedback has been acted on it will go to a copy editor who will suggest stylistic changes, check for logic and consistency, and so forth.
At the same time I will be choosing which plates to put in the book, which images to use on the back cover, writing their descriptions and deciding where to cite them; checking the sources and further reading sections for each chapter and formatting the references; and producing an appendix that lists the scientific names against the vernacular names that I am using in the book. I also need to finalise the acknowledgements section.
As an author, producing a book is a long process that doesn’t end with the actual writing of the manuscript. It’s incredibly satisfying, however, and working with Pelagic on my second book for them has been a great experience. All being well, Birds & Flowers should be out by early winter.
Now, I have three options for the next book that I’m writing….which one to choose…?
As part of our roles as ambassadors of the new conservation organisation Restore (more of which later this year), several of us including Dave Goulson, George McGavin, and myself, are promoting this online petition to get the government to take the issue of neonicotinoid pesticides seriously. Here’s some text from Dave explaining the situation with a link to a petition that you can sign:
“For three years in a row our government has granted farmers special permission to use banned neonicotinoid pesticides on sugar beet. This is contrary to the expert advice of their own Expert Committee on Pesticides, who specifically recommended that permission should not be granted. It also flies in the face of a huge body of scientific evidence showing that these chemicals are phenomenally toxic to all insect life, and that their use on any crop contaminates soils, hedgerow plants, and nearby streams and ponds for years to come. We are in a crisis, with insect populations in freefall. It is about time our government woke up to this, and acted accordingly. This petition https://petition.parliament.uk/petitions/631948 is a necessary means of holding the government to account. Please sign and share, as signing will ensure the issue is debated in Parliament.”
This petition now has more than 15,000 signatures which ensures that it gets a response from the Government. If it reaches 100,000 mark, it will trigger a debate in Parliament. Please sign and promote this important initiative!
For the past year I’ve been keeping a secret from all but a few trusted confidantes: the subject and title of my next book! My publisher – Pelagic – has now announced it on their website and so it’s time to make it public. “Birds & Flowers: An Intimate 50 Million Year Relationship” will be the first book that covers bird pollination in its entirety, going beyond just hummingbirds, sunbirds and honeyeaters, to consider the more than 60 other bird families that interact with flowers, and the tens of thousands of plants that rely on them as pollinators. You can read more about it on Pelagic’s website.
The 3D mock-up of the cover shown above features an illustration by my good friend Stephen Valentine, a very talented artist who you may remember produced this painting of waxwings that Karin bought for my birthday a few years ago. I’m extremely pleased with how Pelagic have incorporated this into the design of the cover.
The book will be available by autumn I hope, if my writing schedule goes to plan!
It’s been a couple of years since I last did a talk or workshop for the Wildlife Trust for Bedfordshire, Cambridgeshire & Northamptonshire. But I’m pleased to say that they’ve invited me back and you can join me tomorrow evening for an online introductory talk about pollinators and pollination in the UK.
The talk starts at 7pm UK time and full details of how to sign up are in the link below:
Note: This is a guest blog post by Guthrie Allen who is a PhD candidate at the University of East Anglia.
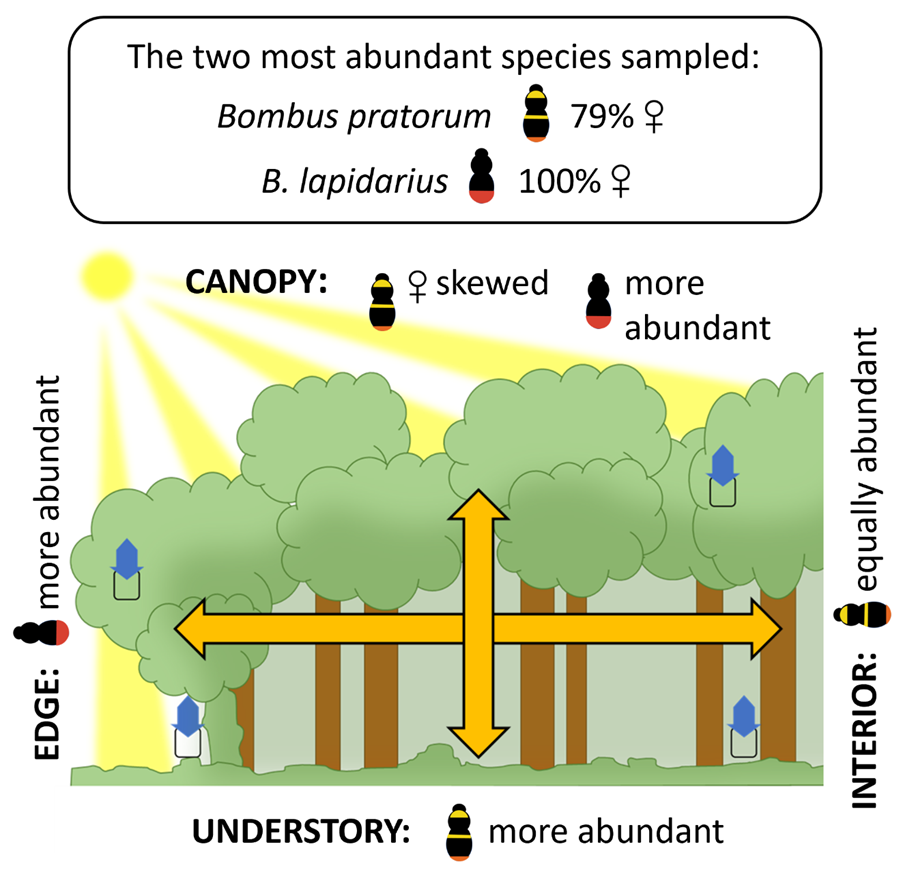
Though often overlooked, woodlands have great potential to support wild bees in farmed landscapes. Both the understory and canopy can provide large quantities of early-season nectar. The pollen of woodland-tree taxa such as Acer can form a large proportion of the early-season diets of bumblebees and solitary bees. Even the pollen from wind-pollinated trees such as Quercus can form a significant part of the diets of several common solitary bee species. Woodlands are also believed to provide ample nesting opportunities. These factors may explain why several studies have found positive correlations between bee abundance and/or diversity and woodland cover at the landscape scale. Despite this, a large-scale European study has shown that interior woodland understories support very few bees compared to exposed woodland edges.
Sampling the canopy – at heights of up to 20 metres – is not easy, but unlike the understory, this habitat remains sun-exposed after canopy closure and could be favourable for bees. In Europe, however, our knowledge of bee activity in the woodland canopy is very limited. Furthermore, whilst canopy-tree taxa have been identified as suitable food sources for many bees, we have little evidence that these trees are used for forage when located within woodlands. To explore the role of the canopy, we trapped bees in late spring in the understory and canopy – at the exposed edges and in the interiors – of 15 woodland sites across an English agricultural landscape.
A significant proportion of bee abundance was found in the canopy, represented by 23 of the 29 total sampled species. Interior canopy activity was much higher when nectar-producing Sycamore (Acer pseudoplatanus) trees were nearby. Communities differed between the canopy and understory: of the seven most common species sampled, three were more active in the understory, while the opposite was true of one species – Bombus lapidarius. Interestingly, we found the sex ratio of the most abundant species – B. pratorum – to be female-skewed in the canopy. For four of these species, and contrary to expectations, we found no evidence that understory activity was any higher at woodland edges compared to woodland interiors.
Further research is needed to understand community differences between the canopy and understory. Nonetheless, our results suggest that both these habitats have a significant role to play in supporting farmland bees. We demonstrate that a diverse bee community has the potential to exploit canopy resources, with nectar-producing trees in woodland interiors likely to provide forage for many bees. And we show the unexpected potential of interior woodland understories to support bee abundance.
Here’s the full reference with a link to the open access paper:
Allen, G. & Davies, R.G. (2022) Canopy sampling reveals hidden potential value of woodland trees for wild bee assemblages. Insect Conservation and Diversity. Available from: https://doi.org/10.1111/icad.12606
Here’s the abstract:
Woodlands can play an important role in supporting bee abundance and diversity in agricultural landscapes. However, in temperate-region studies, the canopy is rarely sampled, and our understanding of its contribution is limited. To explore this, we sampled bees in late spring with blue vane traps in the understory (n = 30) and crowns of mature Quercus robur (n = 35) at the exposed southern edges and in the interiors (ca. 25–75 m from woodland edges) of 15 woodland sites across an English agricultural landscape. A significant proportion of bee abundance and diversity was found in the canopy: canopy-trap catches were estimated to be a third as large as understory-trap catches, and 23 of the 29 sampled species were present in the canopy. Of the seven most common species sampled, four were equally abundant in woodland edge and interior traps; three were more abundant in understory traps, and a single species—Bombus lapidarius—was more abundant in canopy traps. The sex ratio of the most abundant species, B. pratorum, was female-skewed in the canopy. Additionally, the presence of nearby Acer pseudoplatanus trees in flower greatly increased canopy-trap catches in woodland interiors. These results suggest that both the woodland canopy and understory have a significant role to play in supporting farmland bee communities; they indicate the importance of nectar-producing trees in woodland interiors, opening avenues for canopy-based management; and they demonstrate that a diverse bee community has the potential to exploit canopy floral resources.
Within the last decade there’s been a growing awareness of the importance of urban environments for supporting populations of pollinators, especially bees. Indeed, I devoted a whole chapter of my book Pollinators & Pollination: Nature and Society to the topic, though even then I was only able to scratch the surface of the research that’s been done. Since then there’s been some important studies published and this 2020 review by Kath Baldock provides a good starting point for the topic, whilst a recent pre-print by Pietro Maruyama and colleagues emphasises how little we know about pollinators in tropical cities.
One of the most detailed studies of urban solitary bees in a British town was conducted by Muzafar Sirohi when he was a PhD researcher in my department in Northampton. The first paper from that work, documenting the diversity and abundance of bees, came out in 2015, but since then commitments to other projects, plus Muzafar’s return to his university in Pakistan, have meant that we’ve struggled to find the time to publish more. Hopefully that’s changing and the second publication from Muzafar’s thesis is now out, with a third in progress.
This new paper uses a network approach to study the use of flowers by these bees; here’s the reference with a link to a read-only copy of the paper, followed by the abstract.
Biodiversity is declining through human activities and urbanisation is often seen as a particular concern. Urban settings, however, provide diverse microclimatic conditions for plants and pollinating insects, and therefore may be significant habitats for the conservation of solitary and primitively eusocial bees, a major group of pollinators. This study analysed the interactions between these bees and the plants on which they forage, using a network approach. We compared urban habitats (gardens, roadsides, and open vegetation) in a large British town with nearby nature reserves. One native plant Taraxacum officinale (dandelion) was a core generalist species visited in all habitat types. Other core plant species restricted to particular habitats include species of Geranium, Bellis, Crepis, and Ranunculus. Two generalist bee species, Anthophora plumipes and Osmia bicornis were the core visitor species within the networks. The networks were comparatively more nested in urban habitat types than nature areas, suggesting more frequent interactions between generalist and specialist species in urban areas. Network connectance, network level specialisation (H2’ index), and plant generality (network level) were not significantly different in urban and nature areas. However, visitor generality was found to be significantly higher in urban gardens than in nature areas. Careful management of common urban vegetation would be beneficial for supporting urban wild pollinators.
One of the projects with which I’ve been involved over the past few years has been a collaboration with researchers at Imperial College and the Natural History Museum, alongside regional collections in the UK, to assess how museum specimens of bumblebees (Bombus spp.) can be used to look at long-term ecological changes in pollinator populations. The first two papers from that project were published in August but because of my trip to Kenya I’ve only now been able to post about them.
The details of the papers (both of which are open access and free to download) are below, followed by the official press release:
Arce, A., Cantwell-Jones, A., Tansley, M., Barnes, I., Brace, S., Mullin, V., Notton, D., Ollerton, J., Eatough, E., Rhodes, M., Bian, X., Hogan, J., Hunter, T., Jackson, S., Whiffin, A., Blagoderov, V., Broad, G., Judd, S., Kokkini, P., Livermore, L., Dixit, M., Pearse, W. & Gill, R. (2022) Signatures of increasing environmental stress in bumblebee wings over the past century: Insights from museum specimens. Journal of Animal Ecology 00, 1– 13. https://doi.org/10.1111/1365-2656.13788
Mullin, V. E., Stephen, W., Arce, A. N., Nash, W., Raine, C., Notton, D. G., Whiffin, A., Blagderov, V., Gharbi, K., Hogan, J., Hunter, T., Irish, N., Jackson, S., Judd, S., Watkins, C., Haerty, W., Ollerton, J., Brace, S., Gill, R. J., & Barnes, I. (2022). First large-scale quantification study of DNA preservation in insects from natural history collections using genome-wide sequencing. Methods in Ecology and Evolution, 00, 1– 12. https://doi.org/10.1111/2041-210X.13945
OFFICIAL PRESS RELEASE: Museum collections indicate bees increasingly stressed by changes in climate over the past 100 years
• An analysis of bumblebee wings from a network of UK museums shows signs of stress linked to increasingly hotter and wetter conditions. • As well as revealing what is linked to stress in bees in the past, the study can help predict when and where bees will face most stress and potential decline in the future. • Bumblebees and other insect pollinators have faced population declines in recent years. • The researchers have also for the first time used ancient DNA techniques to sequence bumblebee genomes dating back over 100 years. Scientists from Imperial College London and the Natural History Museum today published two concurrent papers analysing UK bumblebee populations.
The first investigated the morphology (body shapes) of bee specimens dating back to 1900. Using digital images, the group first investigated the asymmetry in bumblebee wings as an indicator of stress. High asymmetry (very differently shaped right and left wings) indicates the bees experienced stress during development – an external factor that affected their normal growth.
Studying four UK bumblebee species, the group found evidence for stress getting higher as the century progressed from its lowest point around 1925. Further analysis showed that each bee species displayed a consistently higher proxy of stress in the latter half of the century.
Learning from the past to predict the future By taking the climate conditions during the year of collection – namely annual mean temperature and annual rainfall – the team found that in hotter and wetter years bees showed higher wing asymmetry. The study is published today in the Journal of Animal Ecology.
Author Aoife Cantwell-Jones, from the Department of Life Sciences (Silwood Park) at Imperial, said: “By using a proxy of stress visible on the bee’s external anatomy and caused by stress during development just days or weeks before, we can look to more accurately track factors placing populations under pressure through historic space and time.”
Author Dr Andres Arce, now at the University of Suffolk, stated: “Our goal is to better understand responses to specific environmental factors and learn from the past to predict the future. We hope to be able to forecast where and when bumblebees will be most at risk and target effective conservation action.”
Senior author Dr Richard Gill, from the Department of Life Sciences (Silwood Park) at Imperial, said: “With hotter and wetter conditions predicted to place bumblebees under higher stress, the fact these conditions will become more frequent under climate change means bumblebees may be in for a rough time over the 21st century.”
DNA from a single leg As well as measuring the wing shapes of bees, in a second parallel study the team successfully sequenced the genomes of over a hundred bumblebee museum specimens dating back more than 130 years. In a pioneering advance, ancient DNA methods typically used for studying woolly mammoths and ancient humans, were for the first time used on an insect population.
Scientists from the Natural History Museum and the Earlham Institute quantified DNA preservation using just a single bee leg from each of the bees studied to create a baseline genome for each of the four species.
From these developments, published today in Methods in Ecology & Evolution, the researchers can now look to determine how the reported stress may lead to genetic diversity loss.
In conjunction with providing a new reference genome, the team will now use this data to study how bee genomes have changed over time, gaining an understanding of how whole populations have adapted – or not – to changing environments.
The value of museum collections Focusing on bumblebee collections, the team worked with curators from the Natural History Museum London, National Museums Scotland, Oxford University Museum of Natural History, World Museum Liverpool, and Tullie House Museum Carlisle.
Author Dr Victoria Mullin, from the Natural History Museum, said: “Museum insect collections offer an unparalleled opportunity to directly study how the genomes of populations and species have been affected by environmental changes through time. However, they are a finite resource and understanding how best to utilise them for genetic studies is important.”
Senior author Professor Ian Barnes, from the Natural History Museum, said: “One of the main problems with museum collections is that the quality of DNA can be very variable, making it difficult to predict which type of analyses we should do. We now have a much better idea about DNA preservation in insect collections, which is a massive boost to our ongoing work to understand the history and future of insect populations.”
Dr Gill concluded: “These studies showcase the value of leveraging museums specimens to go back in time and unlock the past’s secrets. But what we have done is just the beginning, and by continuing our work with these vital public collections and collaborating with curators we can only discover more. All this work was part of a Natural Environment Research Council-funded project and could not have been achieved without the commitment, hard work, and diligence of the museum curators, and our other collaborators”.
Rail travel is my favourite form of transport and always has been. I like its slower pace and the fact that on a long journey you can sit back, read or work, and watch the landscape unfold. Not only that but it’s one of the most environmentally friendly types of mass transport. So when I was asked to be one of the lead authors on a report outlining how the European Rail Network can support biodiversity along its 230,000 km length, I was happy to be involved! And that’s one of the main projects that I’ve been working on over the past 12 months, during which I’ve researched the literature, written and revised drafts, and learned a huge amount about the ecology of the rail network!
The report, which was formally released yesterday, was commissioned by the International Union of Railways (Union Internationale des Chemins de Fer or UIC), founded in 1922 as the global industry body for rail transport. The writing and desk top research was led by the UK Centre for Ecology and Hydrology (CEH).
That’s not the end of my involvement, however: the next stage is a technical guide and I’m already starting to work on that. I will report back once it’s complete.
As kids, my friends and I did a lot of digging. We always seemed to be burrowing into slopes or excavating trenches, pretending to be archaeologists or treasure hunters. Indeed, there was a lot of ground treasure to be found in the part of Sunderland where I grew up. The area has a long history of pottery and glass making, and ship building, and the remnants of these industries could be uncovered every time we stuck a spade in the earth. Over time I developed my own small museum of interesting, unearthed fragments, including bits of hand-painted ceramics, glass bottles, and unidentifiable metal shards, alongside various animal bones I’d excavated. My parents quietly indulged this interest, and my muck-streaked face and clothes, even if they didn’t quite understand what I was doing.
Aged about 10, my first encounter with a bumblebee nest was during one such dig. On the waste ground behind a large advertising hoarding, we began digging into a low, grass-covered mound and accidentally excavated what was probably a small nest of Buff-tailed Bumblebees (Bombus terrestris). I can recall being fascinated by the waxy, odd shaped cells and by the sticky fluid that some of them were leaking. Being an adventurous sort of child I tasted the liquid: it was sweet and sticky, and that was my first encounter with bumblebee “honey”.
I’m going to leave those quotation marks in place because if you do an online search for “do bumblebees make honey?” you generally find that the answer is “no, only honey bees make honey”.
Now, defining honey as something made by honey bee strikes me as a circular argument at best. And it also neglects the “honey” made by meliponine bees that is central to the culture of stingless bee keeping by indigenous groups in Central and South America, and the long tradition pre-colonial tradition of honey hunting by Aboriginal Australians. So if we widen our definition of “honey” as being the nectar*-derived fluid stored in the nests of social bees, then Apis honey bees, stingless bees and bumblebees must all, by logic, make honey. And likewise there’s wasps in the genus Brachygastra from Central and South America that are referred to as “honey wasps” because, well, I’m sure you can work it out!
But this is where things become a little trickier, because turning nectar* into honey involves some complex evaporation and enzymatic activity, so that the resulting fluid is more concentrated and dominated by the sugars glucose and fructose. Although analysis of honey bee honey is commonplace, and there’s been some research conducted on the honey of stingless bees, I don’t know of any studies that have compared Bombus honey with that of other bees, or with what is stored in the nests of honey wasps**. If I’ve missed anything, please do comment and let me know, but this strikes me as an area of research demanding some attention.
So do bumblebees make honey? That very much depends on our definitions, but I’m happy to accept that they do because “honey” is not a single thing: it’s an insect-derived substance that can take a range of forms but serves the same broad purpose of feeding the colony. And although insects have probably been producing it for millions of years, I think I’ve known the answer to the question for almost 50 of them…
UPDATE: A couple of people have commented on social media that there are legal definitions of “honey” as a foodstuff. Here’s the definition according to UK law***:
“the natural sweet substance produced by Apis mellifera bees from the nectar of plants or from secretions of living parts of plants or excretions of plant-sucking insects on the living parts of plants which the bees collect, transform by combining with specific substances of their own, deposit, dehydrate, store and leave in honeycombs to ripen and mature”
So, legally, we can’t call anything that isn’t made by Apis mellifera “honey”, at least from a foodstuffs regulation perspective. But that’s clearly different to what we have been discussing above, which is about a biological definition of honey.
It’s also interesting to look at the compositional requirements of honey as a foodstuff (presented in Schedule one of that document, if you follow the link above). The lower limit for moisture content is 20%. Now if you consider that most nectar in flowers has a sugar content of between about 20% and 50%, clearly there’s been a lot of evaporative work done by the bees to reduce the amount of water in the honey. I would love to know how bumblebee (and other insect) “honey” compares to this: do they put the same kind of effort into evaporating the water from the stored nectar? Given that the purpose of reducing the water content is to prevent fermentation by yeasts when it’s stored for a long time, and that there are bumblebee species which have colonies that are active for more than one year, I imagine that at least some species in some parts of their range may employ similar tactics.
Thanks to everyone who has been commenting and discussing the topic. It never ceases to amaze me how much we still do not understand about some fundamental aspects of the natural history of familiar species!
*And honeydew to a greater or lesser extent.
**I’m going to ignore honey pot ants for now as this is complex enough as it is and they don’t store the “honey” in nest cells.
***From what I can gather definitions in other countries are similar.
Following on from my recent blog post about biological crusts, I was intrigued by the patterns formed by these lichens on the clay tiles capping the brick gate columns of our local cemetery. It looks as though they have been created by successive waves of growth, but I may be wrong about that. Any lichen experts out there who can tell me what’s going on?
I think the species is Xanthoria parietina, but again I’m happy to be corrected. Below is a cropped close-up from a slightly different angle.