Earlier this year I was invited by the editor of British Wildlife magazine to write a piece for their Changing Perspectives section about how odd Europe is when it comes to bird pollination. It’s based on one of the chapters in my book Birds & Flowers: An Intimate 50 Million Year Relationship.
If you subscribe to the magazine, it will appear in the August issue, though I’m happy to send a PDF to anyone who doesn’t subscribe (or has not read the book) – use the Contact Page. The main accompanying photograph is by one of my former students, Lisa King, who kindly allowed me to use it.
I’m a long-standing fan of his YouTube video channel which Joey describes as “A Low-Brow, Crass Approach to Plant Ecology & Evolution as muttered by a Misanthropic Chicago Italian.”
It was a lot of fun to talk flowers and pollinators with him and although I tried to keep my swearing to a minimum, if you know Joey and his work, you know what you’re in for, so be warned! It’s not for the easily offended.
We had sound issues at a couple of points and note that at 54:20 I made an error, and said “hummingbirds” a couple of times when I meant “sunbirds”. Put it down to a lack of coffee that morning….
Walking into Kunming Institute of Botany yesterday morning, I passed a young guy who was carrying what I initially thought was a species of Orobanchaceae. I’ve a long-standing interest in the pollination ecology of these intriguing parasitic plants, so I stopped to have a chat. Turns out they were in fact orchids! Specifically, they were specimens of Gastrodia elata, one of the “potato orchids“, so named because those fat tubers are edible. They are widely used in South China – where they are known as Tianma, 天麻 – both as a food and medicinally. The tubers are eaten before the flowers are produced, and originally they were collected from the wild. But in the 1960s a Chinese botanist named Xuan Zhou discovered how to cultivate them and they are now grown in specialist nurseries. A fascinating account of the life of Xuan Zhou – “The Father of Gastrodia” – was published in the journal Plant Diversity last year, shortly after he died.
These orchids do not produce green leaves or stems, therefore they cannot photosynthesise. Instead, they gain all of their energy from a parasitic symbiotic relationship with a fungus – they are what is termed “myco-heterotrophic“. Most myco-heterotrophic plants have evolved from ancestors that were involved in mutualistic mycorrhizal relationships with fungi, in which the plant provides sugars to the fungus in return for mineral nutrients and water. In the case of Gastrodia elata, the fungus concerned is the non-mycorrhizal, wood-rotting Armillaria mellea. In the west we know this as Honey Fungus, a disease of trees and shrubs and the bane of many a gardener. This is also edible, incidentally, but best dried before cooking (and some have an intolerance to it, so take care).
I tweeted the photograph in a short thread just after taking it, and Stewart Nicol pointed me to a study of the orchid’s floral biology and pollination ecology in Japan by Naoto Sugiura. Turns out that, at least in the population which Naoto studied, the plant produces no nectar and deceives its pollinators, which are small bees, into visiting the flowers.
That’s why I’ve used the phrase “doubly-parasitic*” in the title of this post – the plant, it appears, parasitically exploits both the fungus from which it gains energy and the pollinators that ensure its reproduction. It’s (almost, but not quite) the flip side of “double mutualism” in which species provide two benefits for one another, e.g. the same bird is both a pollinator and a seed disperser of a particular plant, a phenomenon that I discussed in my recent book Birds & Flowers: An Intimate 50 Million Year Relationship.
But note the question mark in the title of this post. There’s an enormous amount that we don’t know about these myco-heterotrophic interactions and how they remain stable over the evolutionary history of the plant and the fungus. In order to be considered a parasite, by definition, an organism must have a negative impact on the reproductive fitness of its host. Do these orchids negatively impact either the fungus or the bees that pollinate it? As yet we don’t know. And I was intrigued by this comment from a 2005 review of ‘The evolutionary ecology of myco-heterotrophy‘ by Martin Bidartondo:
“no successful plant lineage would be expected to cheat both mycorrhizal fungi (by failing to provide photosynthates) and deceive insect pollinators (by failing to provide nectar or other rewards) due to the evolutionary instability inherent to specializing on two lineages.”
At first glance it appears that Gastrodia elata is a plant lineage that has done just that, though I’d like to see more work carried out on this system. Specifically, are all populations of the orchid bee pollinated and are all rewardless? And does this orchid really provide no benefit to the fungus, perhaps by synthesising secondary compounds that protect the Armillaria from infection by bacteria or being eaten by invertebrates. So many questions to be answered about this fascinating species interaction!
*With thanks to my wife Karin Blak for inspiring that phrase.
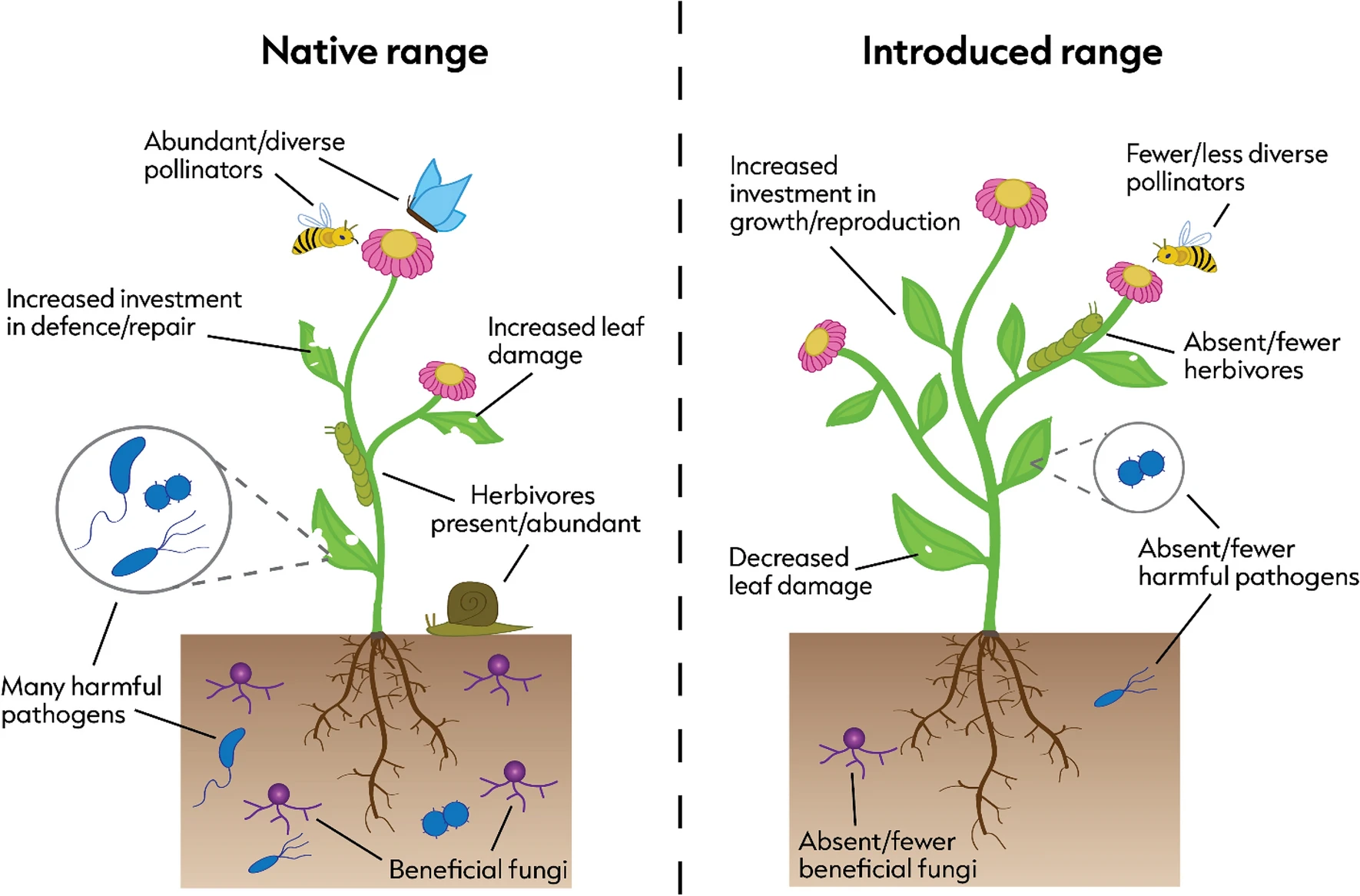
As I’ve previously discussed on the blog, when species are moved to a different part of the world they lose many of the ‘enemies’ – such as predators, herbivores and pathogens – that would normally keep their populations in check. This can have implications for the likelihood of a species becoming invasive, and it’s called the Enemy Release Hypothesis (ERH) and has been well studied. Less well researched is the flip side of the ERH, the Missed Mutualist Hypothesis (MMH), in which species lose their ‘friends’, such as pollinators, seed dispersers, symbiotic fungi, and so forth. It’s a topic I’ve worked on with my colleagues at the University of New South Wales, principally Angela Moles and her former PhD student Zoe Xirocostas.
Another paper from Zoe’s PhD work has just been published and in it she carried out a comparison of European plants that have been transported to Australia, and asked whether they had fewer pollinators in their new range. It turns out that they do!
Here’s the full reference with a link to the paper, which is open access:
Many studies seeking to understand the success of biological invasions focus on species’ escape from negative interactions, such as damage from herbivores, pathogens, or predators in their introduced range (enemy release). However, much less work has been done to assess the possibility that introduced species might shed mutualists such as pollinators, seed dispersers, and mycorrhizae when they are transported to a new range. We ran a cross-continental field study and found that plants were being visited by 2.6 times more potential pollinators with 1.8 times greater richness in their native range than in their introduced range. Understanding both the positive and negative consequences of introduction to a new range can help us predict, monitor, and manage future invasion events.
The UK media has fueled something of a moral panic over the last couple of years, in relation to the Yellow-legged Hornet (Vespa velutina) which has become established as an invasive species in Europe. It also looks likely to become established in Britain and Ireland, where beekeepers have claimed that it poses “a severe threat to pollinators“. The only study that I know of that’s tested this idea in Britain – by Thomas O’Shea-Wheller, Juliet Osborne, et al. – suggests that the impact on bumblebees, at least, is not as great as feared.
In Asia, where the species originates, they’ve lived with this hornet for centuries and learned to exploit it. On a visit to a recent farmer’s market near Kunming we encountered a local man selling bottles of adult hornets steeped in alcohol, to be used as a liniment. It’s rubbed on arthritic joints and (apparently) soothes the pain.
The guy who was selling the bottles of embrocation had several hornet’s nests on display:
Later, on a trip to Lijiang I also spotted a hornet’s nest on a building, not the usual place you expect to see one:
The other use for hornets is as food – the larvae are apparently quite delicious and very nutritious. This is from a different market and is a different species:
Later on the Lijiang trip we visited a farm that was part of a Yi community, one of the local ethnic minorities. They keep the indigenous honey bees (Apis cerana) in these small hives:
The honey bees pollinate an early-flowering local cherry variety that farmers grow in small orchards. The fruit is extremely small but also extraordinarily sweet:
These ones are past their best though still edible:
Much fresher cherries were being sold in farmer’s markets and at the roadside:
Is it too early to talk about Christmas? Not if you’re interested in pollinators and pollination! The mid-winter festival has featured quite a number of times on my blog over the years, especially in relation to the iconic plants that represent this time of year in Northern Europe, and what one might describe as the ‘cultural biodiversity‘ of Christmas. The final plant that I included in that last post was the poinsettia (Euphorbia pulcherrima) – this is how I described it:
In many ways this is an unusual plant to have such a strong cultural association with Christmas: it’s a mildly toxic species of spurge from tropical Mexico that was introduced to North America in the 19th century, then subsequently to Europe. However its festive connotations date back to the earliest period of Spanish colonisation in the 16th century, so it’s older than some…other Christmasy traditions…
Just occasionally one sees a bird-pollinated tree planted in a city. The most common in my experience are various banksias in Australia, and the Royal Poinciana (from Madagascar) and the African Tulip Tree in the urban tropics and subtropics elsewhere in the world. I’ve also occasionally encountered large specimens of Poinsettia: when they are given free rein they are a much more impressive plant than their Christmas cousins. The vivid red bracts that surround the clusters of flowers suggest that they may be hummingbird-pollinated in their native Central America, but as far as I know their pollination ecology has not been studied.
Here at the Kunming Botanic Garden there’s several quite large specimens of poinsettia that, as I write, are in full flower, their red bracts a signal to pollinators that can be seen for quite a distance. However we’ve not seen any of the local sunbirds or white-eyes visit the flowers, and, as I said in the book, as far as I know the pollination ecology of poinsettia has never been studied in the wild. Close inspection of the flowers in the garden revealed that almost all of the nectaries had at least one nectar-collecting ant sticking out from it, their prominent backsides a deterrent to the Asian Honey Bees (Apis cerana) that also wanted a piece of the action.
Based on the position of the nectaries in relation to the stamens, if the plant is hummingbird-pollinated then the pollen is likely to end up under the chin of the bird. That’s certainly been described in other plant-bird pollination systems. But it does not have to be birds that move the pollen around – red flowers are also associated with other kinds of pollinators, for example butterflies and beetles. But until someone in Mexico does the necessary field work, we’ll just have to speculate.
In the image above, the three glass tubes on the right contain different cultivated varieties of soybean (Glycine max), one of the world’s most important (and controversial) legume crops. The tube on the left contains the seeds of what is considered the wild ancestor of soybean (Glycine soja). Archaeological evidence suggests that by about 5,000 years ago farmers in several areas of Asia had artificially selected varieties that had much bigger seeds than the wild type. I like this image because it’s a great demonstration of the power of humans to positively influence the food that we eat – and the power of those seeds to affect us via our diets and farming methods.
Of course it’s not just soybean that’s been subject to this sort of artificial selection – we’re encountering many other species and varieties of beans at the various farmer’s markets we’re visiting here in Yunnan:
Many of these beans require pollinators such as bees to initiate or at least enhance the crop, as we found in our recent study of soybean as part of the SURPASS2 project.
I took the main image last week when I had a tour of the Germplasm Bank at the Kunming Institute of Botany, a really impressive facility that stores both seeds in deep freeze and living plants in tissue culture. It’s one of the ways in which we can help to conserve the genetic diversity of wild and cultivated plants. Here are a few more photographs from that visit:
Dr Jie Cai, the manager of the seed bank, kindly hosted the tour and introduced the facility:
Seeds are stored at about -20C in huge, security-controlled freezers:
Collected seeds are first cleaned, sorted, and then counted – a laborious task that requires patience:
Samples of seeds are also germinated at various points to assess how well they are responding to storage:
Plants with seeds that do not respond well to being stored, such as many orchids and bamboos, are grown in tissue culture:
The building also hosts the genetic sequencing facility, another extremely impressive set of laboratories in which progress of sequencing the Chinese flora can be seen in real time:
During the 2020 lockdown caused by the COVID-19 pandemic, I coordinated an international network of pollination ecologists who used standardised methods to collect data in their gardens. I blogged about it at the time – see here and here for instance – and also put up a post when the data paper from that work was published.
Several research groups are now working with that huge data set and interrogating it for answers to a wide range of questions. The first group to actually publish a paper from the data is a largely Chinese set of researchers from the Key Laboratory of Plant Resources, Conservation and Sustainable Utilization, at the South China Botanical Garden in Guangzhou, assisted by Kit Prendergast and myself.
In this paper we’ve considered how robust these plant-pollinator networks are to simulated extinctions of species, and how this is affected by the elevation, latitude, and plant species diversity of the network.
Here’s the full reference with a link to the study:
If you can’t access it and need a PDF, please send me a request via my Contact page.
Here’s the abstract:
Plant-pollinator interactions play a vital role in the maintenance of biodiversity and ecosystem function. Geographical variation in environmental factors can influence the diversity of pollinators and thus, affect the structure of pollination networks. Given the current global climate change, understanding the variation of pollination network structure along environmental gradients is vital to predict how global change will affect the ecological interaction processes. Here, we used a global plant-pollinator interaction data collection by the same sampling method at the same period to explore the effects of elevation, latitude, and plant richness on the structure and robustness of pollination networks. We analyzed a total of 87 networks of plant-pollinator interactions on 47 sites from 14 countries. We conducted a piecewise structural equation model to examine the direct and indirect effects of elevation, latitude, and plant richness on the network robustness and analyzed the function of network structure in elucidating the relationship between robustness and these gradients. We found that plant richness had both positive effects on robustness under random and specialist-first scenarios. Elevation, latitude, and plant richness affected network connectance and modularity, and ultimately affected network robustness which were mediated by nestedness under specialist-first and random scenarios, and by connectance under the generalist-first scenario. This study reveals the indirect effects of elevation, latitude, and plant richness on pollination network robustness were mediated by nestedness or connectance depended on the order of species extinctions, implying that communities with different pollination network structures can resist different extinction scenarios.
In this delightful book, [Ollerton] describes the ways that birds and flowers interact. As in his previous book, Pollinators & Pollination: Nature and Society, [he] takes a deeply personal approach to the subject. He combines anecdotes from his research travels around the world, to mountains of Kenya and Tanzania, the Andes of Peru, Brazil, and Nepal, among other places, with his contributions to, and masterful knowledge of, the recent literature…
The review is free to read and download from Journal of Pollination Ecology. I’m so glad that people are enjoying the book – if you’ve bought or borrowed a copy, please do leave a comment and let me know what you think.
A message from Dr Debora Drucker, WorldFAIR Agricultural Biodiversity Case Study Lead:
Registration is open to our contribution to the WorldFAIR webinar series – “Reusing Plant-Pollinator Datasets: a Global Perspective with Guidelines and Recommendations inspired by Pilot Studies from Africa, the Americas and Europe”.
We will present results from Deliverables 10.2 & 10.3, with focus on our pilot studies:
Drucker, D., Salim, J. A., Poelen, J., Soares, F. M., Gonzalez-Vaquero, R. A., Ollerton, J., Devoto, M., Rünzel, M., Robinson, D., Kasina, M., Taliga, C., Parr, C., Cox-Foster, D., Hill, E., Maues, M. M., Saraiva, A. M., Agostini, K., Carvalheiro, L. G., Bergamo, P., Varassin, I.; Alves, D. A., Marques, B., Tinoco, F. C., Rech, A. R., Cardona-Duque, J., Idárraga, M., Agudelo-Zapata, M. C., Marentes Herrera, E. Trekels, M. (2024). WorldFAIR (D10.2) Agricultural Biodiversity Standards, Best Practices and Guidelines Recommendations (Version 1). Zenodo. https://doi.org/10.5281/zenodo.10666593
Drucker, D. P., Salim, J. A., Poelen, J., Soares, F. M., Gonzalez-Vaquero, R. A., Devoto, M., Ollerton, J., Kasina, M., Carvalheiro, L. G., Bergamo, P. J., Alves, D. A., Varassin, I., Tinoco, F. C., Rünzel, M., Robinson, D., Cardona-Duque, J., Idárraga, M., Agudelo-Zapata, M. C., Marentes Herrera, E., Taliga, C., Parr, C.S., Cox-Foster, D., Hill, E., Maués, M.M. Agostini, K. Rech, A.R., Saraiva, A. (2024). WorldFAIR (D10.3) Agricultural biodiversity FAIR data assessment rubrics (Version 1). Zenodo. https://doi.org/10.5281/zenodo.10719265
We reserved a good amount of time for Q&A – I hope to see you there and have a nice discussion!