In these turbulent times it’s hard to know where to focus one’s gaze. Do we concentrate on Ukraine? Greenland? Venezuela? Sudan? China? Russia? The Middle East? The rise of the far right and religious fundamentalism? Cyber security? Global organised crime? If it’s confusing and worrying for the average person, imagine what it’s like for national security services who are charged with assessing and responding to such threats.
It is increasingly recognised that national security in the 21st century extends beyond military threats to encompass food systems, economic resilience, public health, and the stability of critical ecological infrastructure. Which is why it’s no surprise to learn that the UK’s national security organisations – MI5 and MI6 – have just released a report titled Global biodiversity loss, ecosystem collapse and national security: A national security assessment.
The global attack on nature is threatening the UK’s national security, government intelligence chiefs have warned, as the increasingly likely collapse of vitally important natural systems would bring mass migration, food shortages and price rises, and global disorder.
This framing explicitly treats biodiversity loss not as an environmental side issue, but as a systemic risk multiplier capable of amplifying existing geopolitical, economic, and social stresses. I have emboldened two words in that quote in order to emphasise that this report is very much about the state of the world, not just the state of my home country. In an interconnected global food and trade system, ecological collapse in one region rapidly propagates elsewhere through markets, migration, and political instability. What happens globally has implications locally; not just food security from imports, but “geopolitical instability, economic insecurity, conflict, migration and increased inter-state competition for resources”, to quote the report.
Where does pollination fit into this? As far as I know, pollination has never been singled out in security analyses, yet it underpins many of the very food systems, rural economies, and ecosystem functions upon which national resilience depends. In my book Pollinators & Pollination: Nature and Society I mention “food security” about ten times, as I firmly believe that loss of pollinators is a serious issue to food supply chains. The report similarly states that:
UK food production is vulnerable to ecosystem degradation and collapse. Biodiversity loss, alongside climate change, is amongst the biggest medium to long term threat to domestic food production – through depleted soils, loss of pollinators, drought and flood conditions.
But I would go further and state that loss of pollination by insects and vertebrates poses a national security threat that extends far beyond just their role in food production.
Let me explain why I believe this.
The Global biodiversity loss report focuses on six different parts of the world (and seven ecosystems) that it considers “critical ecosystems…at risk of collapsing”. One of those areas – the coral reefs of Southeast Asia – is not directly dependent upon pollinators to support its long-term functioning. Two areas – the boreal forests of Canada and Russia – are dominated mainly (though not exclusively) by wind-pollinated trees, such as conifers and birches. The other four ecosystems, however, have a dependence on pollinators that ranges from significant to enormous. These are the Mangroves of Southeast Asia and the Himalayas (both significant) and the Amazon Rainforest and Congo Basin (both enormous).
What do I mean here by words like “significant” and “enormous”? What is my measure? What I mean is the number and proportion of flowering plants—particularly dominant species, often trees—that underpin most ecosystem functions, such as photosynthesis and carbon storage, and that rely to some extent on pollinators to reproduce.
In high elevation areas such as the Himalayas, I know from experience that it’s common for there to be a mixture of wind and animal pollinated species in communities. Similarly, mangrove species include some which are wind pollinated – see this review for example. In other words, the long-term population stability of Himalayan woodland and Southeast Asian mangrove forests is, in large part, dependent on the pollinators that those ecosystems support. If those pollinators were lost, in the long term (decades to centuries) wind-pollinated trees would dominate and biodiversity would significantly decline.
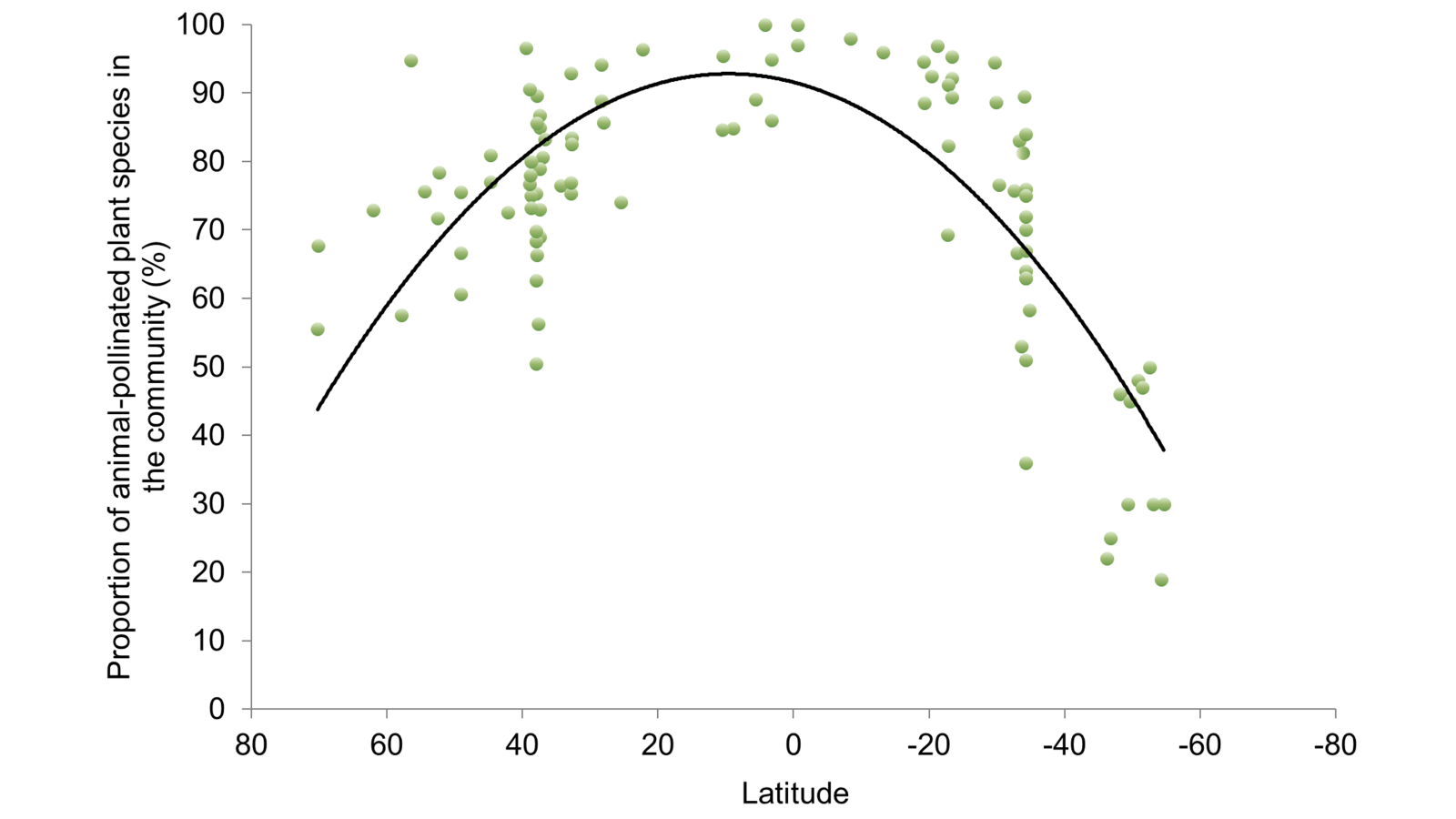
The situation in the forests of tropical South America and west Africa is rather different. Not only is there a much greater diversity of plant species in these ecosystems, but in these largely rainforest regions, often all of them are animal pollinated, as we showed in this paper and which is reflected in the graph above, which comes from my book. Lose the pollinators and we lose the long-term viability of ecosystems that provide regionally- and globally-vital functions.
Ultimately, if we are to protect pollinator communities, and the ecosystem functions and services they provide, we need to take their conservation more seriously than we do at the moment. Framed this way, pollinator conservation becomes a form of preventive security investment, analogous to maintaining flood defences or safeguarding energy and cyber infrastructure. The European Union’s Pollinator Initiative and the projects that it supports, including Butterfly and ProPollSoil in which I’m involved, is a good example. Likewise, there are policy movements appearing in China, as I recently reported. But biodiversity conservation is a global issue, as the security services report makes clear, and that applies to pollinators.
There will no doubt be sceptics out there who think that I am over-playing the importance of pollinators and pollination. That’s fine, it’s good to have these debates. Pollination is not a national security issue in the narrow, traditional sense of defence against hostile actors. But in the 21st-century security landscape, where threats are systemic, slow-burning, and ecologically grounded, pollination loss clearly qualifies as a strategic risk to national stability and resilience.
In that respect, I believe that the question is not whether pollination is a national security issue—but whether national security thinking has yet fully adapted to the biological foundations on which societies depend.
Christmas is a time of expectations, when we assume that familiar traditions will work as they always have. Some traditions are associated with plants such as holly and mistletoe. In ecology, we make similar assumptions, one of the most enduring of which is that plants introduced to new parts of the world often thrive because they leave their natural enemies behind. The thinking goes that leaves that are tough, bitter, or spiky (e.g. holly) or chemically well-defended (e.g. mistletoe), should only strengthen that advantage.
But as with many things we take for granted at this time of year, this intuition does not always hold.
Over the past few years I’ve been part of a large international collaboration led by Zoe Xirocostas (as part of her PhD work) and Angela Moles (see here and here for example) asking deceptively straightforward questions such as: can we predict which introduced plants will experience this “enemy release” based on how well defended their leaves are? In other words, if a plant is tough, hairy, bitter, toxic, or nutritionally poor, does that make it more or less likely to escape herbivores when it turns up somewhere new?
The latest paper from that work has just been published in the journal Oecologia. In it we focused on 16 widespread plant species that are native to Europe but have become established in Australia. For each species, we measured how much leaf damage they suffered in their home range and how much in their introduced range. Our earlier work had already shown that, on average, these plants suffer far less herbivory in Australia – clear evidence of enemy release.
The next step was to ask why.
So we measured an unusually wide range of leaf traits linked to palatability and defence. Some were physical: leaf toughness, thickness, dry matter content, hairiness. Others were chemical: nitrogen content, lipids, phenolics, cyanogenic compounds – the kinds of substances that make leaves bitter, toxic, or difficult to digest. We also combined these into overall “chemical”, “physical”, and “total defence” scores.
If plant defence theory works the way we often assume, at least some of these traits should have explained why certain species escape enemies more than others.
They didn’t.
Across all of these traits – individually and in combination – we found no relationship at all with the strength of enemy release. Plants with heavily defended leaves did not escape herbivores any more than plants with relatively soft, palatable foliage. Equally striking, leaf defences didn’t even predict how much damage plants suffered overall.
This was a genuinely surprising result, especially given how central plant defence traits are to ecological thinking. It suggests that when plants are introduced to new regions, the details of leaf chemistry and toughness may matter far less than we assumed.
One possibility is that context trumps traits. Herbivory may depend more on which insects happen to be present locally, how closely related the newcomer is to native plants, or how chemically novel it is relative to its neighbours, rather than on how well defended it is in absolute terms. Another is that many defensive traits serve multiple roles – coping with drought, heat, or pathogens – and are not just about deterring herbivores. As an example, consider cactus spines, that not only defend a plant against hungry herbivores, but also provide shade for the stem, collect water from fog or dew, and insulate the plant against extreme hot or cold temperatures.
From a practical perspective, our findings have implications for managing invasive species. Biological control programmes – such as the ones that are targeting Japanese Knotweed and Himalayan Balsam in the UK – often assume that poorly defended plants are easier targets. Our results suggest that defence traits alone may not be a reliable guide.
What makes this study particularly powerful is its scale. The same species were measured using the same protocols in different countries; fieldwork was coordinated across multiple climates and ecosystems; statistical analysis drew on shared expertise. This kind of international cooperation can be slow, complex, and occasionally frustrating – but it is exactly what is needed if we want to move beyond neat ideas tested in a single place.
Ecology advances not just through clever hypotheses, but by discovering when our intuitions fail. In this case, a widely held assumption about plant defences and enemy release simply didn’t hold up. That doesn’t close the book on the question – but it does force us to turn the page and rethink what really determines success and failure when species move around the globe.
If you are interested in the full study, here’s the reference with a link to the paper, which is open access:
At a time when the UK’s wildlife is under increasing pressure, the everyday spaces we manage—especially gardens—are becoming ever more important. Although interest in wildlife-friendly gardening has grown enormously in recent years, the evidence behind different approaches is not always clear. Well-meaning interventions can be highly effective, but some can miss the mark without a grounding in sound ecological knowledge.
That’s exactly why the Wildlife Gardening Virtual Symposium has become such a valuable annual event. It brings together researchers, practitioners, and anyone involved in managing green spaces to explore what the science is actually telling us about creating gardens that support biodiversity.
This year’s programme, chaired by Hafsah Hafeji of the Wildlife Gardening Forum, features four invited talks covering ponds, pollinators, fungi, and urban mammals, along with an update on emerging policies and projects shaping the wildlife-gardening landscape.
2026 Speaker Programme
Fragments of Paradise: Garden Ponds as Wildlife Habitat Dr Mike Jeffries – Northumbria University
Gardening for Pollinators: It’s About More Than Just Flowers! Prof Jeff Ollerton – University of Northampton & Kunming Institute of Botany
How Fungi Make Gardens Flourish Dr Jassy Drakulic – Royal Horticultural Society
Recording Wild Mammals in Urban Spaces: A Multidecadal Study David Wembridge – People’s Trust for Endangered Species
Whether you’re involved in ecology, horticulture, landscaping, consultancy, education, or simply interested in the future of wildlife in our gardens, the symposium offers a concise way to catch up on current evidence and emerging thinking.
The ProPollSoil project officially got underway on 1st October, and this week (16th–20th November) our consortium gathered in Freising, Germany, for the kick-off meeting hosted by the Technical University of Munich. It was an inspiring start: dozens of experts from across Europe coming together to explore two big questions: How does the health of our soils shape the fate of pollinators? And how do pollinators influence soil health?
Most people think of pollination as something that happens in the air or on flowers, but for many species the story begins underground. Thousands of bees, hoverflies, beetles and wasps depend on soil to nest, overwinter, or complete parts of their life cycle. For example, around half of the solitary bee species in Britain and Ireland are what we term “ground nesting” and make their nests in different types of soil. Yet soil conditions—structure, temperature, contaminants, farming practices—are changing rapidly. As part of the EU’s Mission Soil programme, ProPollSoil aims to understand these hidden links so we can better protect the pollinators that support our food systems and ecosystems.
The project brings together specialists in entomology, soil science, ecology, modelling, agriculture, economics, and communication, forming a truly interdisciplinary team. Through desk-based reviews, fieldwork, lab experiments, monitoring and advanced modelling techniques, we’ll be investigating how soil influences pollinator survival and what we can do to improve it.
ProPollSoil is built around six key goals, including identifying the soil conditions that help pollinators thrive, testing innovative ways to monitor soil-dependent species, evaluating how different land-use and farming practices affect pollinators, and developing practical soil-management solutions—from reduced tillage to cleaner soils—that can slow or reverse their decline.
My own role mainly focuses on understanding the state of our current knowledge of the biology and ecology of soil-dependent pollinators and their interactions with soils, other invertebrates, and plants. I’ll also be working on integrating information about pollinators’ soil dependencies into the European Atlas of Plant-Pollinator Associations (EuroAPPA), part of the related Butterfly Project, whose kick-off meeting I documented on the blog earlier this year.
Together, these efforts will help build a clearer, more complete understanding of how life belowground supports life aboveground. It’s an exciting journey, and we’re only just getting started!
My sincere thanks to all of the ProPollSoil consortium members whose passion and expertise made for a stimulating few days in Germany. And a special shout-out for the team from Poland who brought with them some delicious, PropPollSoil branded sweets:
Last week I returned from a 14 day visit to China to colleagues at the Kunming Institute of Botany in Yunnan, part of a three-year commitment to working there that I documented on the blog last year, starting here. Some of my recent trip involved a long weekend in the city of Nantong, just north of Shanghai, where I was an invited speaker at the International Pollinator Insect Biology and Pollination Symposium. During a full day of talks from researchers and practitioners, via the excellent simultaneous interpretation service provided by the organisers, we learned about recent developments in the world of Chinese honey bees and wild pollinators. There were also international guest speakers from Australia, Argentina, and the UK, in person and online.
First of all, a number of speakers commented on the growing realisation in China that the value of crop pollination services by honey bees (both the native Asian Apis cerana and the European A. mellifera) far outweighs the value of the hive products such as honey, wax and royal jelly – see this from the 2021 study by Shibonage K Mashilingi and colleagues:
The total economic value of pollination amounted to US$ 106.08 billion in 2010, representing 19.12% of the total production value of Chinese agriculture
In comparison, the global honey market was valued at just US$ 9.01 billion in 2022. That such an understanding of the much greater economic value of pollinators to agriculture was relatively slow in coming is perhaps not surprising – it’s easier to weigh a physical product than it is to assess the contribution of bees and other insects to an apple harvest, for instance. But this awareness is a crucial step towards understanding the many reasons why pollinators need protection.
Which leads me to my next point: there was considerable political interest in the conference and in the topic more broadly. The meeting opened with almost an hour of introductory remarks by high-ranking Chinese officials, including the Vice Mayor of the regional government, the Vice President of the Chinese Academy of Agricultural Sciences, and the Secretary General of the Ministry of Agriculture and Rural Affairs of China. All of them commented on the importance of pollination to both crops and wild plants, and the need to reduce the amount pesticides being used in Chinese agriculture. I can’t recall ever being in a pollination symposium in any other country where there was such a political presence. I think that it says a lot about the Chinese willingness to translate science and technology into government policy and actions.
At the end of the opening session I had the chance to talk briefly with Liu Jian, former Vice Minister of Agriculture and Rural Affairs of China. Via an interpreter we agreed on the importance of pesticide reduction for protecting pollinators, a theme he had emphasised strongly in his talk, and I presented him with a copy of my book Pollinators & Pollination: Nature and Society:
Following the opening addresses there was a talk by the President of the Apicultural Science Association of China, Prof. Peng Wenjun, who gave us “An overview of the development of China’s bee pollination industry”. He described pollinators as the “invisible pillar” of agriculture, which is a wonderful phrase, and set out a strategy for greater integration of government policies, science, and technological innovation in order to support both managed and wild pollinators.
The first set of talks ended about 6pm, then it was back to the hotel for a quick dinner, before returning to the venue for a set of 15 shorter, but no less excellent, talks by postgraduate and postdoctoral researchers. This over-ran slightly and finally drew to a close at about 10pm, signalling the end of a very long, but very stimulating, day.
The following morning we were up early for a tour of some local agricultural facilities, including a high-tech glasshouse demonstration project and a loquat orchard that included trees which are thought to be around 300 years old. The thing that links these two contrasting agricultural systems is the requirement for managed pollinators to produce a crop: bumblebees (Bombus spp.) in the case of glasshouse tomatoes and the Asian honey bee (Apis cerana) for the winter-flowering loquat. Here are some photographs from that trip:
My sincere thanks to the organisers of the symposium for the invitation to speak and to my colleagues Zong-Xin Ren, Scarlett Howard, Yuansheng Fu, and Carlos Matallana-Puerto for their companionship on the trip. I’m grateful also to our personal translator-guides Yang and Gao who surprised us at the airport and made us feel so welcome:
When it comes to the statistical analysis of data, I know my limits. Maths was never my strong point at school or university, and my approach has always been to keep analyses as simple and straightforward as possible*, or to rely on colleagues with fancier statistical chops to do the heavy lifting. I wish that were not the case – I wish I had a brain that was more number-focused than it is. But I don’t and I’ve learned to live with it, to play to my actual strengths as a scientist, and to collaborate with others who can bring different skills to the party.
In theory, the development of the R platform was supposed to make life easier for those of us who wanted to analyse complex data sets. But actually having to script, from scratch, the code to carry out even simple analyses always seemed to me to be a step backwards from the push-button days of SPSS or Minitab. Yes, I get that R is incredibly powerful and flexible and blah blah. But it still involves a heavy time commitment and an aptitude for writing code that many of us struggle with.
Recently, however, things have changed. I find myself carrying out complex statistical analyses that would have stumped me 12 months ago. Not only that, but I now understand those tests on a much deeper level than I ever did before. I also feel much more confident in the interpretation of the outputs from the tests I’m running, and their limitations.
Why the over night change? ChatGPT.
More precisely, I’m using ChatGPT to help me decide which analytical approaches are best for the data that I have, getting it to help me to write the R script to carry out the tests, and then (crucially) it’s advising me on the interpretation of the statistical output and suggesting future steps.
Let me give you an example. I’ve just submitted a manuscript to a journal which describes the results from an experiment that had confounded me for years, which is why I’d not published the work previously. Following discussions with some colleagues in China I realised that my framing of the work was wrong (by coincidence, a topic that Jeremy Fox has recently discussed over on the Dynamic Ecology blog). However, there was still a contradiction in two of the sets of results that I could not resolve: they should have been telling me the same thing but they were not. When I queried ChaptGPT on this it suggested that I model the data taking into account the fact that I had missing data – missingness in statistical jargon. When I did – bingo! – the results made sense: the absence of some data in my experimental treatments had systematically biased the results. It all made perfect sense.
Now, I could have talked this over with a statistician or a more statistically-minded ecologist colleague. But scientists are busy people and I did not want to impose on someone’s limited time. Or rather multiple someone’s limited times, because I know from past experience that when you ask folks these sorts of questions you can get different advice depending upon their own experiences, training, or preferred flavours of statistical analysis. By treating ChatGPT as a collaborator I can get an objective answer to my data questions, written in a way that I can understand. That last point is key because for all of us with specific expertise it’s sometimes difficult to translate our knowledge into broadly interpretable language.
How can I know that ChatGPT is giving me reliable statistical advice? It certainly didn’t give me accurate information about Erasmus Darwin a couple of years ago (a story, incidentally, that I included in my recent book Birds & Flowers: An Intimate 50 Million Year Relationship). But since then, the reliability and accuracy of ChatGPT has improved considerably and when I’ve checked the information it’s given about analyses it was usually accurate as far as I can gauge. In one case, however, it completely missed the point of what I was trying to do with another set of data. But of course advice from human collaborators can also be inaccurate – everyone is fallible. So including human (my!) oversight in all of this is important.
I’m certainly not the only one using ChatGPT and other AI platforms in this way – here’s a small sample of some online articles I’ve found on the topic:
I especially like this quote from that last article:
“If I hired a consultant to write the code when I told them what I needed, would that be a problem? Then, what’s the problem in doing stats with an (AI) consultant?”
I can only agree, and again, I emphasise that we need to think of AI as a collaborator as much as a tool.
By now you might be wondering what any of this has got to do with blues musician Robert Johnson**. Well, according to legend, Johnson was initially a fairly mediocre guitar player. His overnight transformation into a brilliant and highly influential guitarist was attributed to him meeting the Devil at a local crossroads and trading his soul for the mastery of his instrument.
Of course the story is nonsense, and Johnson clearly worked hard at his craft, but it feels like a nice metaphor of where we are at the moment: at a crossroads when it comes to how we use AI, with the opportunity for it to turn us, overnight, into experts. Will that come at a profound cost? Are we selling our souls to some kind of digital Devil? AI naysayers claim it will and we are. But as with all technological advancements, from the wheel to the internet, there are both good and bad aspects to it. Only time will tell whether the former outweighs the latter. In the meantime, let’s try to discover how best to use this amazing technology for our own and society’s benefits.
——————————————————————————————————————-
*In fact one of my recent data papers involved no statistical analysis at all because the results were so clear cut: to paraphrase the abstract, when East African lowland honey bees encountered other insects on flower heads, those other insects were displaced 100% of the time. That’s not a result that requires any kind of test.
**Blues is one of my favourite genres of music and this is not the first time it’s featured on the blog. When Karin and I returned to the UK in March 2024 one of the first things that I got involved with was helping to kick-start a regular blues evening featuring professional British and international players. The not-for-profit initiative is called Harborough Blues – check out the website for upcoming gigs.
Image credit: ChatGPT of course, under instruction from me
Earlier this year I received an unexpected invitation from Bloomsbury Publishing to attend a book launch at Philip Mould’s gallery in London. Looking at the details I immediately said yes, because it combined three of my passions: natural history, art, and books! Not only that, but the topic of the book was one very close to my heart – the wildlife of our towns and cities.
Urban Plants is the latest addition to Bloomsbury’s British Wildlife Collection, a stunningly produced series that has set a new benchmark for natural history literature in this country. The author, Trevor Dines, formerly worked for the charity Plantlife, and is a real authority on urban botany. My expectations for this book were very high! So on the day of the book launch, Karin and I trundled down to the capital and spent part of the day at the National Gallery where, among other things, we enjoyed an exhibition by José María Velasco. As well as being a superb documenter of the 19th century landscapes of Mexico, Velasco was also profoundly interested in botany. We’d not planned it that way, but it was a nice coincidence.
The book launch itself was well attended and I found myself catching up with a few familiar faces from the world of British wildlife, and Trevor (whom I’d corresponded with but never met) treated us to a short reading:
I took the opportunity to buy a copy, had a quick chat with Trevor, who kindly signed the book, and then we headed back to catch a train.
So what do I think of Urban Plants?
It’s actually hard to praise the book too much without sounding unnecessarily gushy! But it really is one of the best books that I’ve read for a long time. In part that’s because it stirs deep emotions of me as a child, taking my first faltering steps into the world of natural history on the bomb sites and post-industrial landscapes of my native Sunderland. But it’s more than that: the author writes with elegance and authority on a topic about which he’s deeply passionate, and this comes through on every one of the amply illustrated pages. Trevor should be congratulated on producing a book that will be the go-to reference on the topic for many years to come.
And an important topic it is too: there’s no doubt naturalists who will sneer at the idea of urban botany, but (as the author points out) for many people in this country, the plants that they see every day in their home towns are almost their only connection to wildlife. For that reason alone it’s a subject to be taken seriously, and if a book like this can inspire more people to take a closer look at the plants with which we share our streets, roofs and walls, so much the better.
So do yourself a favour and take a walk with Trevor through the complex ecology and botany of built-up British landscapes. I learned a lot from Urban Plants and I highly recommend it as an addition to anyone’s Christmas list.
The Golden Lotus (Musella lasiocarpa) is one of China’s most iconic plants — a striking member of the banana family (Musaceae) that seems to bloom forever. Its brilliant yellow, lotus-like bracts have long made it a favourite of subtropical gardeners, though it also has utility as a food and fibre crop, and is associated with Chinese Buddhism. As you can see above it often features stylistically in Chinese temples, and in my visits to Yunnan we frequently encounter it during fieldwork on farms, planted to support terraced fields:
But despite its fame, one mystery has lingered for decades: what actually pollinates it?
Until now, Musella was thought to rely mainly on insects, particularly bees, for pollination. That assumption made it something of an outlier within the banana family, where most species are pollinated by birds or bats. But a new study, in which I was involved as part of an international team of predominantly Chinese and Brazilian researchers, has turned that view on its head.
By combining careful field observations with citizen science records, our team found that the Golden Lotus is regularly visited by an impressive diversity of birds — twelve species from five different families. As I documented in my recent book Birds & Flowers: An Intimate 50 Million Year Relationships, many of these visitors, such as bulbuls and sunbirds, are known nectar-feeders, and their behaviour at the flowers suggests that they are acting as effective pollinators. This discovery significantly expands what we know about the pollination ecology of the Golden Lotus, and places it firmly within the broader pattern of bird pollination that characterises much of the banana family.
Interestingly, the plant’s features — large, robust, vividly coloured bracts, abundant accessible nectar, and long-lived blooms — make perfect sense in this new light. These are traits that favour bird pollination rather than the short, concentrated visits typical of bees.
But the significance goes beyond one species. Bird pollination plays a vital, and often overlooked, role in China’s native flora, linking ecosystems from tropical rainforests to mountain valleys. Understanding these relationships is important not only for biodiversity conservation but also for horticulture — helping gardeners and landscape designers to create spaces that attract and sustain pollinators of all kinds.
The Golden Lotus has always been celebrated for its beauty and longevity. Now, we can add another layer to its story: a reminder that even the most familiar plants can still surprise us, and that nature’s partnerships are often more complex — and more colourful — than we imagine.
Here’s the reference with a link to the paper, which is open access:
Pollinators such as wild bees, butterflies, and hoverflies are in trouble worldwide. A major new study, published in Science and led by Gabriella Bishop and other scientists at Wageningen University & Research, shows that the oft-quoted figure of 10% semi-natural habitat in farmland landscapes is far too little to safeguard pollinators. Instead, the evidence points to a need for somewhere between 16% and 37% habitat cover, depending on the type of pollinator, if we are serious about halting declines. Suitable habitats include hedgerows, patches of woodland, species-rich grasslands, and flowering margins, and as a general rule, hoverflies need less of it whilst bumblebees and butterflies require more.
I was fortunate to play a part in this global assessment, contributing an unpublished dataset collected with my former PhD student, Sam Tarrant, who studied plant-pollinator interactions on restored landfill and established grassland sites. Seeing those data joined with dozens of other studies from around the world underlines something we have known for years: no single dataset, however carefully gathered, can give us the whole picture. To really understand what is happening to biodiversity—and to design conservation solutions that work—we need these kinds of global, mega-author syntheses that draw together evidence from many landscapes, taxa, and approaches.
The message from this analysis is stark but hopeful. More habitat means more pollinators, across all groups. Richer habitats with abundant flowers give an additional boost, but the overriding priority must be to increase the sheer area of natural habitat in farmed landscapes. Small-scale fixes like wildflower strips offer short-term benefits, but without enough space they can’t deliver recovery at scale. Long-term, secure habitat creation—on the order of decades, not seasons—is what pollinators, farmers, and ecosystems need.
Although the policy debate in Europe provided the backdrop for this study, the lessons (and the data) are global. Wherever agriculture dominates, the health of pollinator populations—and by extension our food security and biodiversity—depends on our willingness to give these insects the space and quality of habitat they require.
Looking ahead, we need to think bigger and work together. That means more international collaborations, more sharing of data, and more commitment to long-term solutions that transcend borders. The image at the start of this post is from my trip back to China in July this year. I deliberately chose it because, as you’ll see from the map below which is taken from the paper, there was no suitable data available for the study from that country. Or from Africa. Or Australasia. Or from most of tropical South America. That shows that as pollination ecologists we need to coordinate more in advance on these types of syntheses, and maximise the value of the kinds of data that we collect. The main take away from this study, however, is that if we want to reverse the declines in biodiversity, scientists, policymakers, businesses, farmers, and citizens all have a role to play. Pollinators remind us that nature is interconnected and global—our conservation efforts must be, too.
Here’s the full reference with a link to the study:
Biodiversity in human-dominated landscapes is declining, but evidence-based conservation targets to guide international policies for such landscapes are lacking. We present a framework for informing habitat conservation policies based on the enhancement of habitat quantity and quality and define thresholds of habitat quantity at which it becomes effective to also prioritize habitat quality. We applied this framework to insect pollinators, an important part 5 of agroecosystem biodiversity, by synthesizing 59 studies from 19 countries. Given low habitat quality, hoverflies had the lowest threshold at 6% semi-natural habitat cover, followed by solitary bees (16%), bumble bees (18%), and butterflies (37%). These figures represent minimum habitat thresholds in agricultural landscapes, but when habitat quantity is restricted, marked increases in quality are required to reach similar outcomes.