Why do authors write about ‘nature’? What are their motivations and how did they start their writing journey? Do they even recognise this label of ‘nature writer’?
These are just some of the questions I’ll be exploring with two other authors at the Market Harborough Book Festival on Saturday 5th October.
Jack Cornish is author of The Lost Paths, an exploration of the ancient pathways that have criss-crossed England and Wales since prehistoric times, the peoples who made them, and the landscapes through which they currently run. It’s a reminder of ‘just how precious these paths are, and have been, to the human story of this island’. I’ve only just started The Lost Paths, but what I’ve read so far is wonderful. Check out this recent review on The Great Outdoors site.
Ben Masters’ most recent book is The Flitting, an account of the final months of his relationship with his late father, a keen natural historian with a devotion to butterflies, and how they come to share ‘passions, lessons and regrets as they run out of time’. There’s a nice review of The Flitting by Mark Avery on his blog, and I have to agree with him, it’s a lovely book.
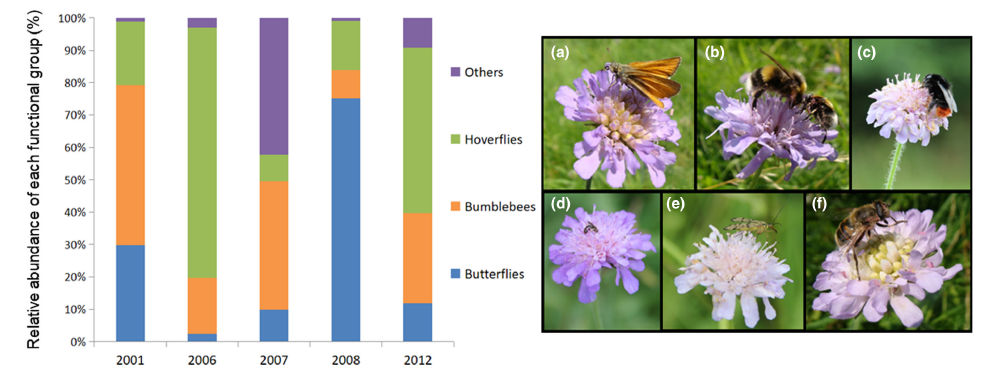
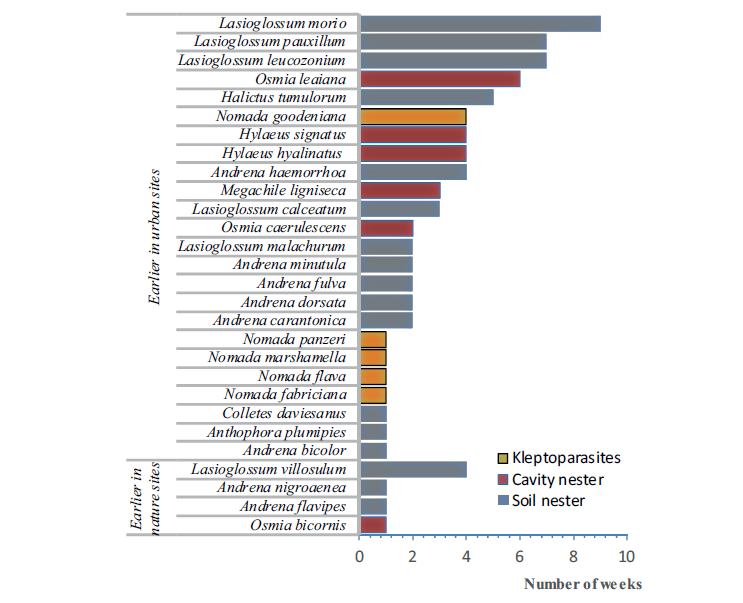
Coincidentally, earlier this year Mark wrote a review of the book that I will be discussing, my recent Birds & Flowers: An Intimate 50 Million Year Relationship, though I may also dip into Pollinators & Pollination: Nature and Society, because there’s at least one thing that unites the three of us as writers: a love of the poet John Clare! Ben discusses him at length in The Flitting, and indeed Clare provided the title of the book. Likewise, Jack name checks Clare in The Lost Paths, and I used the poet as the jumping off point for a couple of explorations of the importance and conservation of bees and other pollinators.
As well as discussing our roles as ‘nature writers’ we’ll be reading extracts from our books and answering audience questions. There will also be an opportunity to buy personally signed copies of our books. We look forward to seeing you there!