We’re coming to the end of our time here in Kenya and we’ve amassed some amazing memories of the wildlife with great views of large mammals such as giraffe, zebra, elephant, hippos, and a variety of antelopes including the ubiquitous dik-diks. In addition, between us we’ve put together a bird list of about 130 species for the site. But as always it’s the smaller things that have fascinated me the most and some of the student project groups have worked with pollinators, ants, and other invertebrates.
One of those fascinations has been the life that exists beneath some of the rocks that are embedded in the red soils of this part of the savanna. Turn them over and you often find the green growths of what are probably cyanobacteria – so-called “blue-green algae” – which are true bacteria and not at all related to algae, even though they also photosynthesise.
The question arises, of course, of how an organism that requires sunlight to survive is able to grow under a stone? Our investigations have shown that they only live under quartz rocks, mainly those that are lighter in colour. Quartz is of course a crystal and it allows a small amount of sunlight to pass through, typically only one or two percent of the sunshine hitting the rocks. There’s also greater humidity under the stones so it’s a relatively more benign place to grow than the savanna, especially in the dry season.
The blog has been quiet over August because Karin and I have been in Kenya for most of the month at the Mpala Research Centre. I’m here teaching on a Tropical Biology Association (TBA) field course, as well as doing some writing. In addition to sharing the adventure, Karin is also writing and acting as unofficial field course therapist!
This is the second TBA field course on which I have taught, the other being in Tanzania back in 2011, and it’s a pleasure to give some time to this remarkable organisation. The model is a very simple one: take 24 students, half from Africa and half from Europe, and embed them in a field work environment for a month, where they learn from one another and from their tutors about ecology and conservation. It’s been hugely successful and TBA alumni now hold senior positions in national conservation departments and NGOs, and universities, across Africa and Europe. Some of the African alumni are also returning to help teach on the field course.
We’re back in Denmark around the 9th September but in the meantime here’s a selection of photographs showing where we are staying and the work that we are doing.
Getting up close with an Acacia species that defends itself by housing colonies of ants in its inflated thorns.
Invasive Prickly Pears (Opuntia spp.) are a growing problem in Kenya, where the cochineal bug has been introduced to help control them.Although there’s an electric fence around the camp site, antelope such as Kudu and Dik Dik are regular visitors. This tent has been our home for most of August. Early in the trip we were confined to it when we both caught COVID. There are worse places to recuperate! The students sorting samples in our open-air classroom, while the White-browed Sparrow Weavers tolerate our intrusionsSpot the snake! The Puff Adder is one of the most deadly snakes in Africa. Fortunately one of the students is an experienced herpetologist and qualified to handle these venomous reptiles.As I write, our TBA students are hard at work on their projects. This is Janeth and Swithin who are looking at competition between honey bees and other pollinators on flowers of this Acacia species.Karin in African ornithologist mode!Examining the Kenya Long-term Exclosure Experiment (KLEE) aimed at understanding the role of mega-herbivores in maintaining savanna biodiversityI’ve donated a copy of my book to the TBA’s Africa library and it’s already inspired some student projects.Sunrise on the savanna
Most of us have at some time stared in fascination at the life contained within the pools that form on rocky shores at low tide. But none of us realized that a whole new class of ecological interaction was taking place!
The 12,000 or so described (and many un-named) seaweeds are incredibly important organisms. Their diverse and abundant photosynthesizing fronds make them one of the main primary producers in coastal seas, creating food and habitat for a huge range of animals. Not only that, but some – the coralline seaweeds – lock up vast amount of CO2 as calcium carbonate and help to create reef systems in the same way as coral.
Although scientists have studied seaweeds for hundreds of years, many aspects of their ecology are still unknown. Their detailed mode of reproduction, for example has only been studied in a small proportion of species.
In a newly published study in the journal Science, French PhD researcher Emma Lavaut and her colleagues have shown that small isopod crustaceans – relatives of woodlice and sea slaters – facilitate the movement of the equivalent of seaweed sperm (termed “spermatia”) from male to female reproductive structures in just the same way that bees and other pollinators move pollen between flowers, so fertilizing female gametes.
Your read that correctly: some seaweeds have pollinators!
It’s an incredible finding! And the implications of this are enormous: Emma and her colleagues have added a whole new branch of life to the examples of sedentary (fixed-place) organisms that require a third party to enable their reproduction. In addition to being a fascinating biological discovery, it has significant environmental and sustainability implications.
Seaweeds are a diverse group of macroalgae that appeared more than one billion years ago, at least 500 million years before the evolution of what we think of as “true” plants, such as the flowering plants, conifers, cycads, ferns and mosses. Sexual reproduction in the brown and green seaweeds, which include kelps, wracks and sea lettuces, involves spermatia that are mobile and use a flagellum to swim through the water to seek out female reproductive structures. However, Emma studied a seaweed, Gracilaria gracilis, which belongs to the Rhodophyta or red seaweeds, and none of the species in this group have these swimming sperm equivalents.
Sexual reproduction in the red seaweeds has therefore always been something of a mystery. Three quarters of species have separate male and female individuals and so they cannot mate with themselves. It was assumed that the gametes were just released into water currents that haphazardly transported them to the female reproductive organs, much as wind pollinated grasses and pine trees release their vast clouds of pollen on land. The authors of this new study, however, point out that most sexual reproduction by these red seaweeds takes place in the relatively still waters of rock pools, a habitat that they mimicked in the laboratory in a series of elegant aquarium experiments.
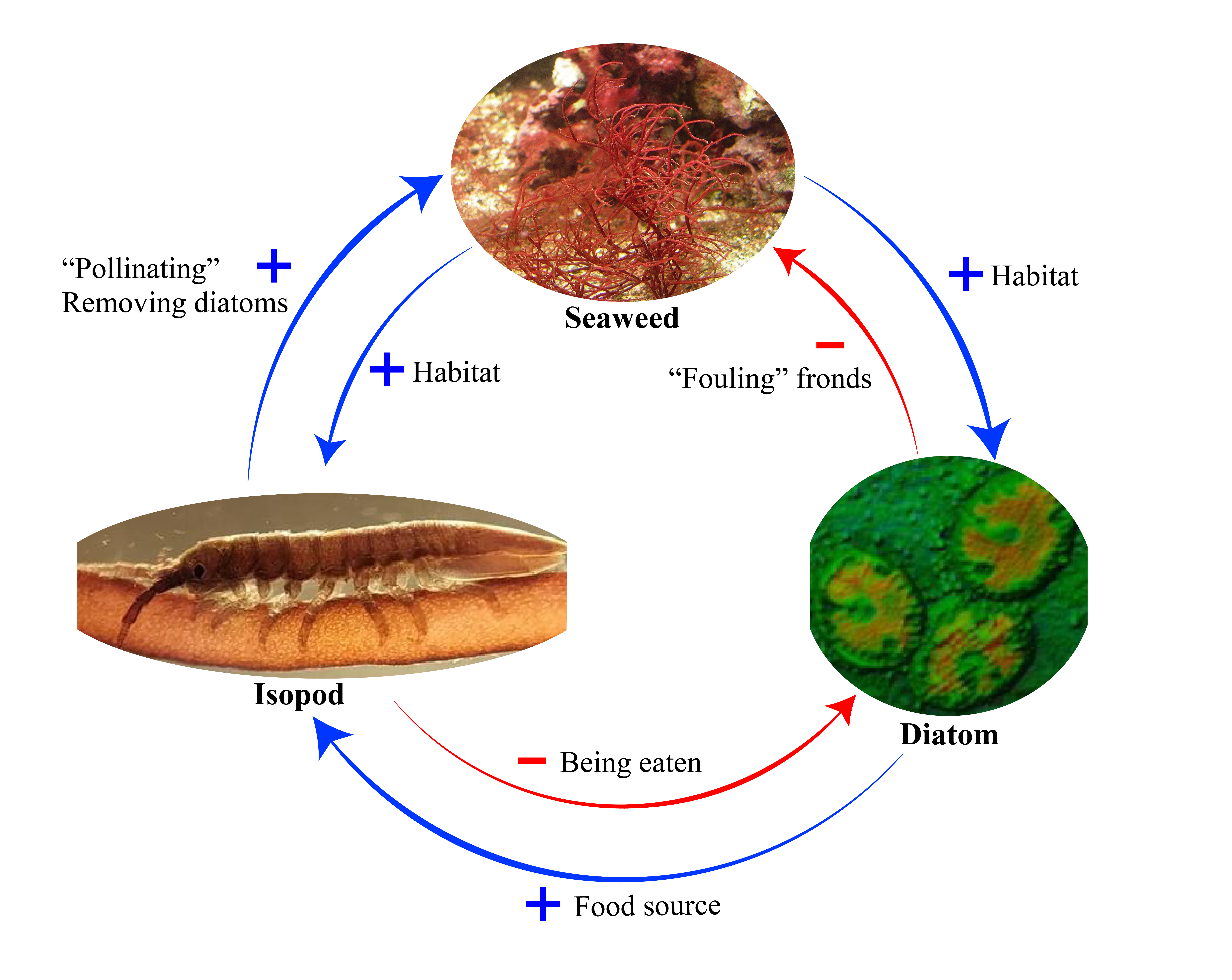
The isopod crustaceans are attracted to the seaweed because they provide a habitat away from predators and a supply of food: they graze on the microalgae that colonise the seaweed’s fronds. Picking up spermatia and moving them between fronds is a side-effect of this activity by the small invertebrates. As you can see from the illustration above, the isopods and the seaweed are engaged in a “double mutualism“: a plus sign (+) indicates a positive effect of one species on another, while a minus sign (-) indicates a negative impact.
What I find especially fascinating about this research is that both the seaweed (Gracilaria gracilis) and the isopod (Idotea balthica) were originally described as species more than 200 years ago. They also have an extremely wide distribution. The isopod is found around the coasts of Europe and down the eastern seaboard of the Americas. The seaweed is pretty much found globally. These are not rare, unusual species, yet the interaction between them has only just been discovered! This is a point that I made in my recent book Pollinators & Pollination: Nature and Society: quite often, species that are well known interact in previously undocumented ways because no one has had the time or inspiration to look closely at them.
Although the idea that small sea creatures might be helping seaweeds to reproduce sounds very fanciful, there is a precedence for this discovery. Back in 2016, in a paper published in Nature Communications, a group of Mexican researchers led by Brigitta van Tussenbroek showed that a species of seagrass is pollinated by a diverse assemblage of small crustaceans and polychaete worms. Seagrasses are flowering plants, not seaweeds, but clearly this type of mutually beneficial relationship can exist between different species in the oceans.
Rhodophyta are the most diverse group of seaweeds, with more than 7,000 known species. They are especially abundant on coastal shores, oceanic habitats that are under huge pressure from infrastructure development, pollution, and climate change. At the same time, these seaweeds are economically important and millions of tonnes of them are collected every year as food, as nutritional and pharmaceutical supplements, and to produce agar. In order to conserve these seaweed populations, we need to better understand their ecology and their environmental requirements.
The work by Emma Lavaut and colleagues suggests that interactions with their “pollinators” may be a critical aspect of this understanding. In the same way that “Save the Bees” has been a rallying call for conserving interactions between species on land, we may soon hear this message echoed in “Save the Isopods”. At the very least, I have to add a new section to the second edition of my book!
Full disclosure: I was one of the reviewers of the original manuscript submitted to Science by Emma and her co-authors. It’s a rare privilege to review a study and think: “Wow! This is a game-changer!” and including this paper it’s happened to me only a handful of times. The editors at Science kindly invited my colleague Dr Zong-Xin Ren and myself to write a Perspective piece about the work and we were delighted to do so.
Image credits: Isopod and diatom images from Lavaut et al (2022). Gracilaria image by Emoody26 at English Wikipedia CC BY 3.0 https://commons.wikimedia.org/w/index.php?curid=3455016. Design by Shijia Wen and Jeff Ollerton.
Over the weekend there was a discussion on Twitter about “beewashing” that was spun out of this tweet by London beekeeper Richard Glassborow. Richard and his colleagues are some of the most responsible beekeepers that I know and they are getting increasingly frustrated by claims from irresponsible companies that keeping a hive of bees in your garden will help to “save the bees”, backed up by spurious claims that “honeybee colonies are dying out”.
The Twitter exchange prompted me to produce the Condescending Wonka meme that you see above because, as I discussed in my recent book Pollinators & Pollination: Nature and Society, pollinator conservation is a really complex area. But there’s no doubt that beekeeping as it’s being widely promoted is not the answer to bee conservation. Let me explain why.
The word “honeybee” does not refer to just one species. It’s most often* applied to bees in the genus Apis, especially the Western Honeybee Apis mellifera, but there are another seven or so Apis species to which the word can be applied. Of those other Apis species, most have never been domesticated and they live as free-living colonies is the various parts of Asia where they evolved. Only Apis cerana is kept in hives, as far as I am aware. The conservation status of most of these other Apis species is unclear but given that they are predominantly forest species, and deforestation is a chronic problem in Asia, we can surmise that some species may be declining. If you want to know more about them the Wikipedia page is a good starting point.
In this short post I just want to consider the Western Honeybee (Apis mellifera). This is a really knotty species to get to grips with because there are multiple subspecies and within subspecies there are various genetic lineages. In addition, the Western Honeybee has been subject to artificial selection for desirable qualities, such as docility, amount of honey produced per hive, and disease resistance, as well as cross-breeding between different subspecies**. The best recent summary of our current understanding of Western Honeybee genetics and conservation is this 2019 review by Fabrice Requier and colleagues, from which I’ve drawn quite a bit of information.
For the purposes of this explaining what’s going on, it’s easiest to think about the species as comprising three “megapopulations”:
Western Honeybees that are managed in hives: For the most part these are not endangered. Britain has as many hives now as it did in the mid-1950s and indeed globally we have more hives than ever (about 90 million hives at the last count). They are found far beyond their natural range and have been introduced into places where they are not native such as the Americas, parts of Asia, and Australia. STATUS: doing just fine.
Western Honeybees that have founded “feral” colonies: These have escaped from hives in countries where they have been introduced and become naturalised. They are doing well, too well in fact: they are a significant conservation issue in places like Australia. STATUS: doing just fine.
Western Honeybees that are living wild in their native range: This is where things become a little muddier. The African populations of the various subspecies seem to be doing well, but more studies are needed to confirm this. In Europe, actually defining what constitutes “wild” honeybees across a region where a lot of selection and hybridization has gone on, probably for thousands of years, is tricky. However there’s no doubt that wild colonies of Apis mellifera are not uncommon in suitable woodland: see this paper about free-living colonies in Ireland by Keith Browne and colleagues, for instance. Note their statement that genetic evidence shows that “the free-living population sampled is largely comprised of pure A. m. mellifera“, i.e. the European Black Honeybee. STATUS: probably doing quite well though more data is needed.
Conclusion: as I said, it’s really complicated and I don’t pretend to have all of the answers, no one does. But what IS clear is that managed Western Honeybees are not declining and keeping yet more hives of them is not going to help us to “Save the Bees”. I’ll leave the last word to Requier et al., whose review I really do recommend: “We argue for the redirection of attention from managed honey bees to the neglected conservation of wild honey bees.” Amen to that.
———————————————————————————————
*The term “honeybee” is sometimes also used for other social bees that produce honey, for example stingless honeybees in the genus Trigona, but there’s no real consensus on what “honey” actually is, and as I’ve argued in another post, bumblebees (Bombus spp.) also produce honey.
If you’ve read my book Pollinators & Pollination: Nature and Society you’ll know that I have a section in the chapter “The shifting fates of pollinators” that deals with the honey bee situation. In that section I bring together the most comprehensive data set so far available on changes in number of hives in Britain. It’s based on a couple of earlier blog posts and if you’ve not read my book take a look at this one first and then this one to give you some context and more information about the sources of the data.
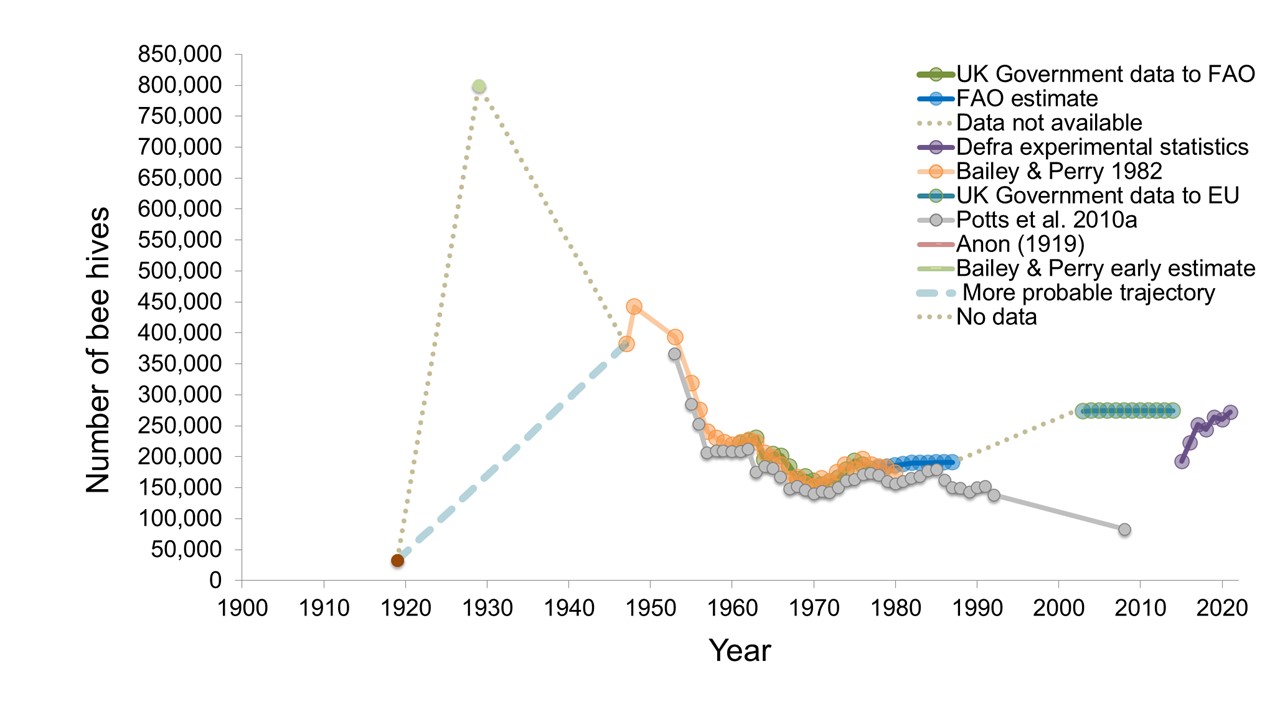
So far this year I have had several requests from people for the original data (which I’m happy to supply) and queries about what it means. So I thought that the time was right to update the graph with the latest official government figures from BeeBase.
The graph above brings the story up to 2021 where the official estimated number of hives is 272,631. That’s an increase of more than 40% since the first BeeBase estimate in 2015.
The take home from this figure is that the current number of honey bee hives in Britain is similar to what it was in the mid-1950s.
So the answer to the question “have honey bees declined in Britain?” is a resounding NO! They are at least as abundant as they were almost 70 years ago. This reflects the global situation where there’s been a substantial increase in hive numbers since the 1960s, as you can see in the figure below.
So if you want to “Save the Bees” or otherwise support pollinators, please focus on the wild, unmanaged species rather than the managed Western Honeybee (Apis mellifera). As always, comments and questions are welcome below or send me a message via my Contact page.
In my recent book Pollinators & Pollination: Nature and Society I discussed the current state of our knowledge of how populations of pollinators have changed over time. Although we have some quite detailed data for particular, often charismatic, species or for certain geographic localities or regions, for most species we know almost nothing. As I wrote in the chapter “The shifting fates of pollinators”:
“For most pollinators we are ‘data deficient’, in other words, we don’t know how their populations are performing. They could be doing well, but they may not be”
This is particularly true for those regions for the world that hold the greatest terrestrial biodiversity: the tropics. For the vast majority of species in the tropics we know precious little about trends in their populations and how their distributions have changed over time in the face of wide-scale land transformation and recent climatic shifts. Filling in some of the gaps in our knowledge of Neotropical pollinator distributions is one of its aims of SURPASS2, a collaboration between South American and UK ecologists, and one of several research and outreach projects with which I’m involved.
In a new study that’s come out of that work, led by Rob Boyd from the UK Centre for Ecology and Hydrology, we’ve used the GBIF database to look at the changing distributions of four important groups of pollinators: bees, hoverflies, leaf-nosed bats and hummingbirds. In particular we were interested in understanding the kinds of biases that come with such publicly available data, and whether recent efforts to add data to GBIF has improved our understanding of trends.
Our overall conclusion is that there are significant limitations and biases inherent in all of these data sets even for groups like hummingbirds which one would imagine are well documented by scientists and bird-watching naturalists. In addition, having more data does not necessarily help matters: it can introduce its own biases.
The paper is open access and feely available; here’s the reference with a link:
Aim Aggregated species occurrence data are increasingly accessible through public databases for the analysis of temporal trends in the geographic distributions of species. However, biases in these data present challenges for statistical inference. We assessed potential biases in data available through GBIF on the occurrences of four flower-visiting taxa: bees (Anthophila), hoverflies (Syrphidae), leaf-nosed bats (Phyllostomidae) and hummingbirds (Trochilidae). We also assessed whether and to what extent data mobilization efforts improved our ability to estimate trends in species’ distributions.
Location The Neotropics.
Methods We used five data-driven heuristics to screen the data for potential geographic, temporal and taxonomic biases. We began with a continental-scale assessment of the data for all four taxa. We then identified two recent data mobilization efforts (2021) that drastically increased the quantity of records of bees collected in Chile available through GBIF. We compared the dataset before and after the addition of these new records in terms of their biases and estimated trends in species’ distributions.
Results We found evidence of potential sampling biases for all taxa. The addition of newly-mobilized records of bees in Chile decreased some biases but introduced others. Despite increasing the quantity of data for bees in Chile sixfold, estimates of trends in species’ distributions derived using the postmobilization dataset were broadly similar to what would have been estimated before their introduction, albeit more precise.
Main conclusions Our results highlight the challenges associated with drawing robust inferences about trends in species’ distributions using publicly available data. Mobilizing historic records will not always enable trend estimation because more data do not necessarily equal less bias. Analysts should carefully assess their data before conducting analyses: this might enable the estimation of more robust trends and help to identify strategies for effective data mobilization. Our study also reinforces the need for targeted monitoring of pollinators worldwide.
——————————————————–
SURPASS2 has been a hugely productive project as you’ll see if you look at the Publications page of the website. There’s much more to come and I’ll report on those research papers as they appear.
Recently I enjoyed chatting with Dr Daniela Scaccabarozzi for the YouTube channel that she runs called Earth To Be. In a wide ranging interview we discussed my recent book, how it came about, some of the things that intrigued me during its research (including a cockroach-pollinated flower!), and the role of people and pollinators in the wider ecosystem. Thanks to Daniela for the invitation to chat! Here’s the link to the interview.
As kids, my friends and I did a lot of digging. We always seemed to be burrowing into slopes or excavating trenches, pretending to be archaeologists or treasure hunters. Indeed, there was a lot of ground treasure to be found in the part of Sunderland where I grew up. The area has a long history of pottery and glass making, and ship building, and the remnants of these industries could be uncovered every time we stuck a spade in the earth. Over time I developed my own small museum of interesting, unearthed fragments, including bits of hand-painted ceramics, glass bottles, and unidentifiable metal shards, alongside various animal bones I’d excavated. My parents quietly indulged this interest, and my muck-streaked face and clothes, even if they didn’t quite understand what I was doing.
Aged about 10, my first encounter with a bumblebee nest was during one such dig. On the waste ground behind a large advertising hoarding, we began digging into a low, grass-covered mound and accidentally excavated what was probably a small nest of Buff-tailed Bumblebees (Bombus terrestris). I can recall being fascinated by the waxy, odd shaped cells and by the sticky fluid that some of them were leaking. Being an adventurous sort of child I tasted the liquid: it was sweet and sticky, and that was my first encounter with bumblebee “honey”.
I’m going to leave those quotation marks in place because if you do an online search for “do bumblebees make honey?” you generally find that the answer is “no, only honey bees make honey”.
Now, defining honey as something made by honey bee strikes me as a circular argument at best. And it also neglects the “honey” made by meliponine bees that is central to the culture of stingless bee keeping by indigenous groups in Central and South America, and the long tradition pre-colonial tradition of honey hunting by Aboriginal Australians. So if we widen our definition of “honey” as being the nectar*-derived fluid stored in the nests of social bees, then Apis honey bees, stingless bees and bumblebees must all, by logic, make honey. And likewise there’s wasps in the genus Brachygastra from Central and South America that are referred to as “honey wasps” because, well, I’m sure you can work it out!
But this is where things become a little trickier, because turning nectar* into honey involves some complex evaporation and enzymatic activity, so that the resulting fluid is more concentrated and dominated by the sugars glucose and fructose. Although analysis of honey bee honey is commonplace, and there’s been some research conducted on the honey of stingless bees, I don’t know of any studies that have compared Bombus honey with that of other bees, or with what is stored in the nests of honey wasps**. If I’ve missed anything, please do comment and let me know, but this strikes me as an area of research demanding some attention.
So do bumblebees make honey? That very much depends on our definitions, but I’m happy to accept that they do because “honey” is not a single thing: it’s an insect-derived substance that can take a range of forms but serves the same broad purpose of feeding the colony. And although insects have probably been producing it for millions of years, I think I’ve known the answer to the question for almost 50 of them…
UPDATE: A couple of people have commented on social media that there are legal definitions of “honey” as a foodstuff. Here’s the definition according to UK law***:
“the natural sweet substance produced by Apis mellifera bees from the nectar of plants or from secretions of living parts of plants or excretions of plant-sucking insects on the living parts of plants which the bees collect, transform by combining with specific substances of their own, deposit, dehydrate, store and leave in honeycombs to ripen and mature”
So, legally, we can’t call anything that isn’t made by Apis mellifera “honey”, at least from a foodstuffs regulation perspective. But that’s clearly different to what we have been discussing above, which is about a biological definition of honey.
It’s also interesting to look at the compositional requirements of honey as a foodstuff (presented in Schedule one of that document, if you follow the link above). The lower limit for moisture content is 20%. Now if you consider that most nectar in flowers has a sugar content of between about 20% and 50%, clearly there’s been a lot of evaporative work done by the bees to reduce the amount of water in the honey. I would love to know how bumblebee (and other insect) “honey” compares to this: do they put the same kind of effort into evaporating the water from the stored nectar? Given that the purpose of reducing the water content is to prevent fermentation by yeasts when it’s stored for a long time, and that there are bumblebee species which have colonies that are active for more than one year, I imagine that at least some species in some parts of their range may employ similar tactics.
Thanks to everyone who has been commenting and discussing the topic. It never ceases to amaze me how much we still do not understand about some fundamental aspects of the natural history of familiar species!
*And honeydew to a greater or lesser extent.
**I’m going to ignore honey pot ants for now as this is complex enough as it is and they don’t store the “honey” in nest cells.
***From what I can gather definitions in other countries are similar.
Clearly that’s a very subjective question and everyone has their own view on which books about a particular subject they would recommend! So coming up with a list of just five for the Shepherd book recommendation site was not easy. My list features authors such as Brenda Z. Guiberson, Megan Lloyd, Steven Falk, Dave Goulson, Mike Shanahan and Stephen L. Buchmann, which will hopefully inspire you to read some of these books.
If you think that I’ve missed your favourite from the list, please do comment below. And if you’re an author, consider signing up for Shepherd and curating your own list, they’ve been really helpful and it’s a useful service for readers and authors.
Of all of the “classical” pollination syndromes, flowers that are hawkmoth pollinated have one of the highest levels of predictability. If a flower is pale in colour, opens at night, is highly scented, and possesses a long tube at the bottom of which is a supply of nectar, there’s a very high likelihood that it’s pollinated by long-tongued hawkmoths (Sphingidae).
Fast forward 160 years and we now know that pollination syndromes are more complex than 19th and early 20th century scientists imagined – see my recent book Pollinators & Pollination: Nature and Society for a discussion of this topic. That’s not surprising because, as I point out, we probably have data on the interactions between plants and their pollinators for only about 10% of the estimated 352,000 species of flowering plants. There’s still much to be discovered!
As an example of how our understanding of specialised flower-hawkmoth interactions is developing, consider this recent study that I’ve just published with my Brazilian colleague Felipe Amorim and other collaborators. In it we have shown that, contrary to expectations, a species of Apocynaceae (Schubertia grandiflora) with a relatively short floral tube can specialise on hawkmoths with much longer tongues than we might predict.
The full reference with a link to the study is shown below, followed by the abstract. If you would like a PDF, please drop me a line via my Contact page:
Since Darwin, very long and narrow floral tubes have been known to represent the main floral morphological feature for specialized long-tongued hawkmoth pollination. However, specialization may be driven by other contrivances instead of floral tube morphology. Asclepiads are plants with a complex floral morphology where primary hawkmoth pollination had never been described. We detailed here the intricate pollination mechanism of the South American asclepiad Schubertia grandiflora, where functional specialization on long-tongued hawkmoth pollinators occurs despite the short floral tube of this species. We studied two plant populations in the Brazilian Cerrado and recorded floral visitors using different approaches, such as light-trapped hawkmoths for pollen analysis, direct field observations, and IR motion-activated cameras. Finally, using a community-level approach we applied an ecological network analysis to identify the realized pollinator niche of S. grandiflora among the available niches in the pollinator community. Throughout a period of 17 years, long-tongued hawkmoths were consistently recorded as the main floral visitors and the only effective pollinators of S. grandiflora. Flowers rely on highly modified corona and gynostegium, and enlarged nectar chambers, to drive visitors and pollination mechanism. Despite its relative short-tube, network analysis placed S. grandiflora in the module including exclusively long-tongued hawkmoth pollinators and the most phenotypically specialized sphingophilous plants in the community. These results represent the first example of functional specialization in long-tongued hawkmoths in an asclepiad species. However, this specialization is uncoupled from the long floral tubes historically associated with the sphingophily syndrome.