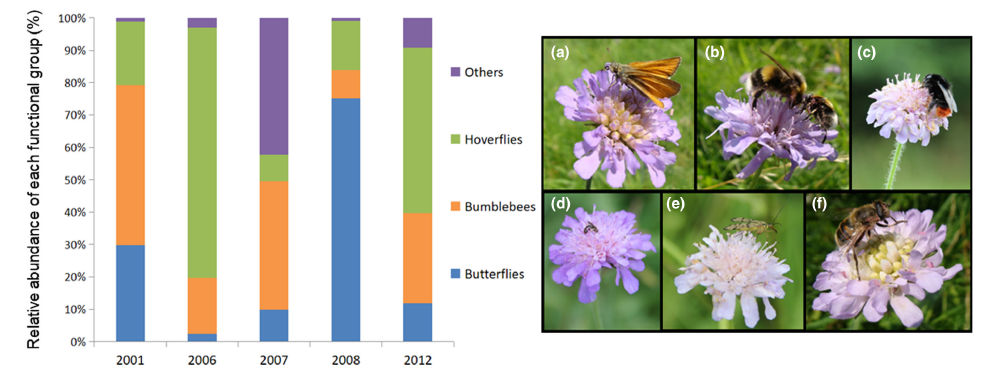
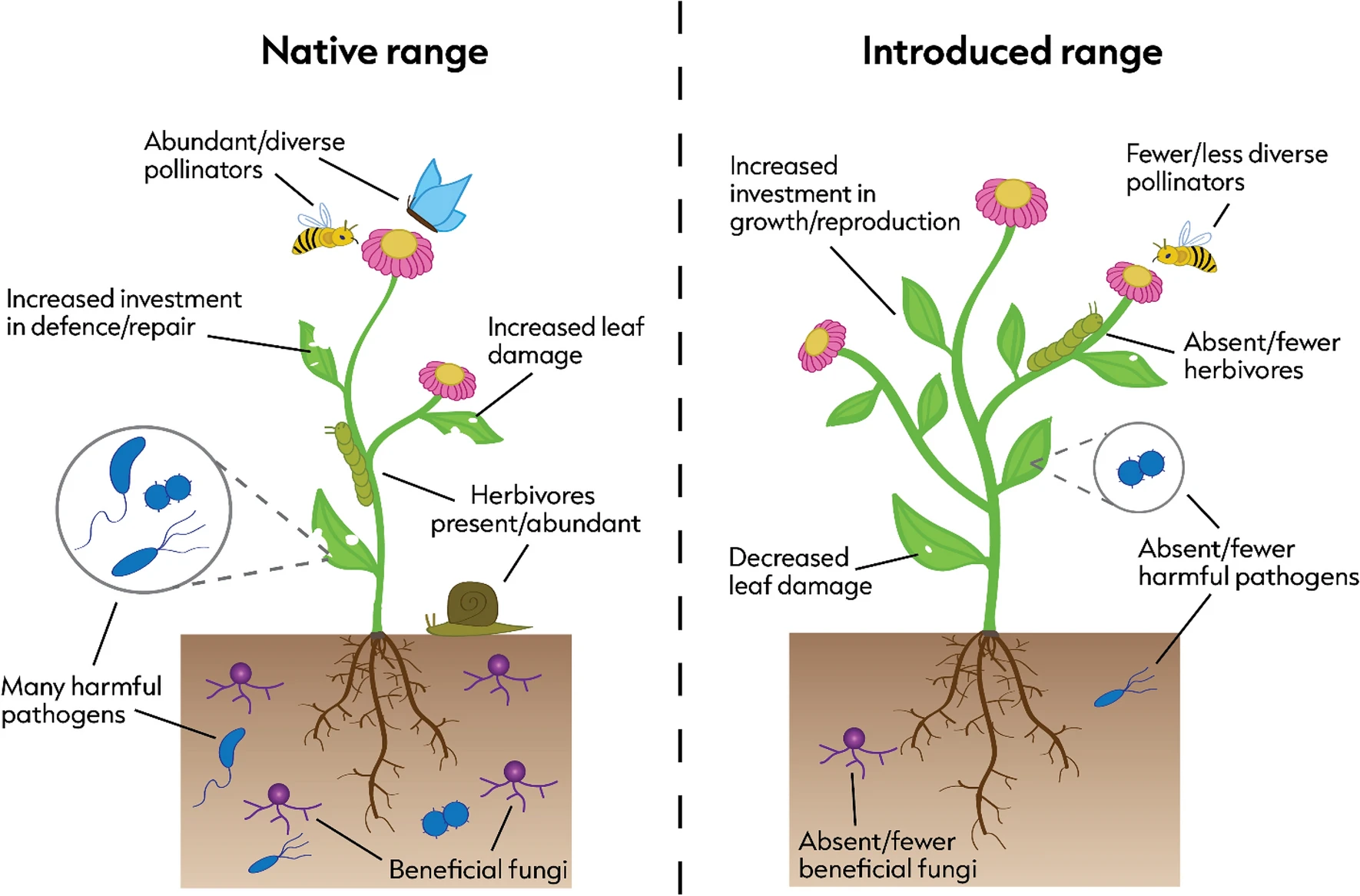
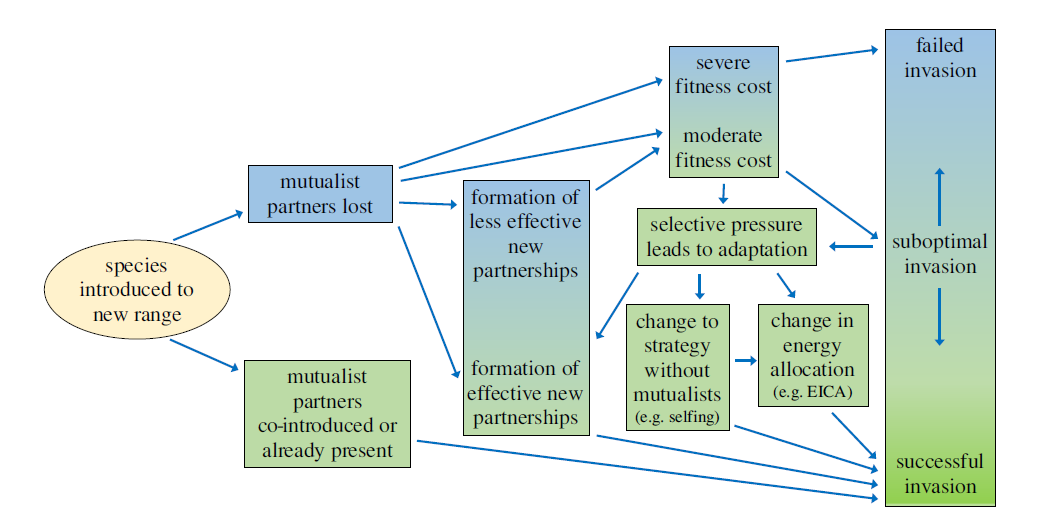
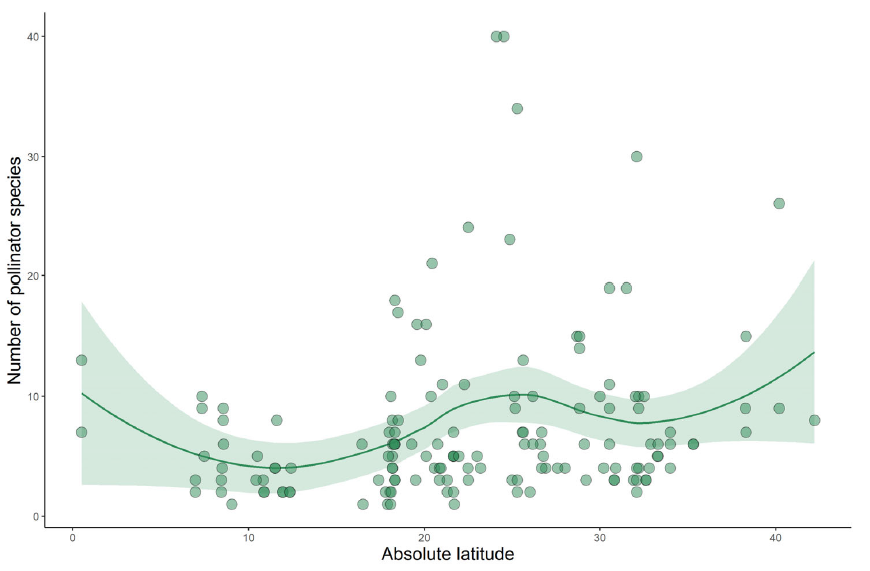
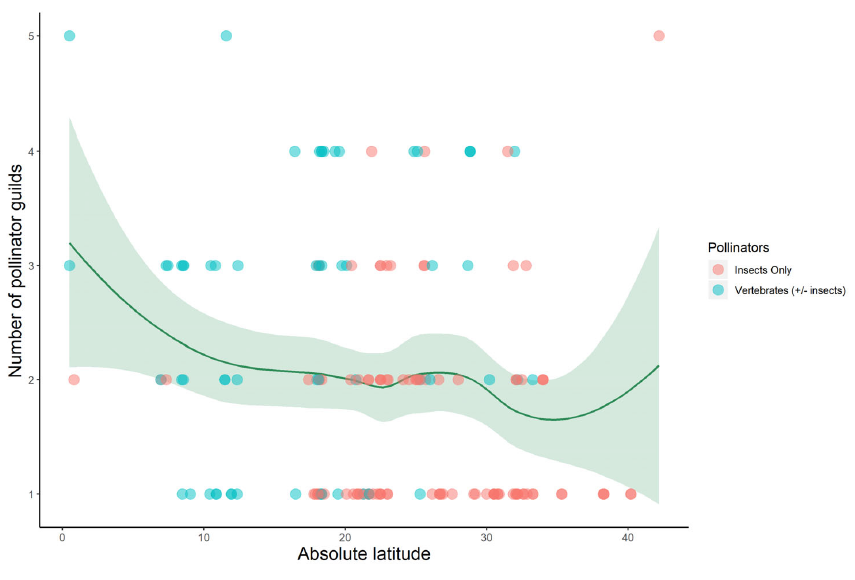
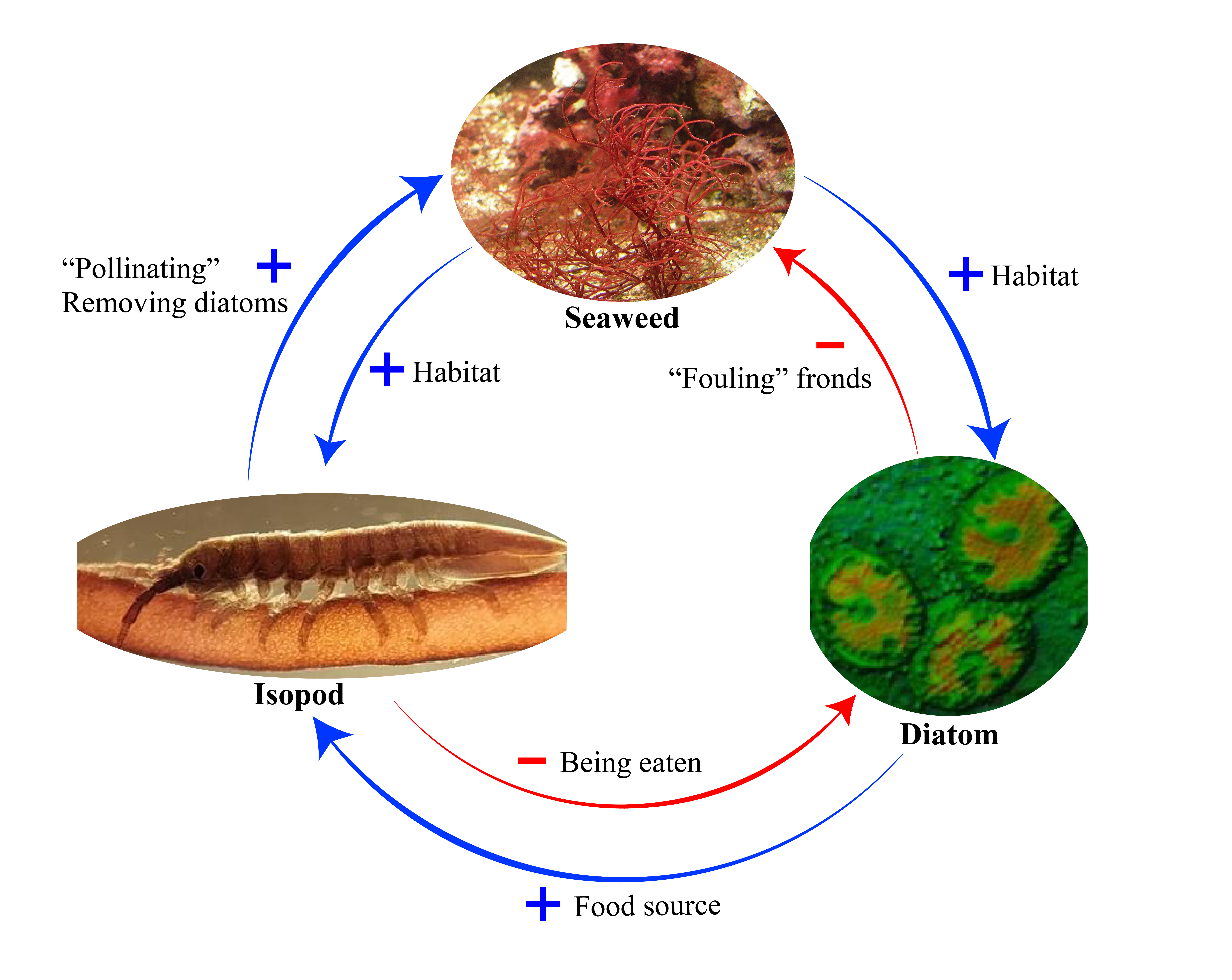
When we think of pollination, we often picture bees buzzing around flowers or butterflies flitting from bloom to bloom. This relationship between plants and pollinators is one of the most well-known interactions in nature. But insect pollination didn’t begin with the colorful flowers we see today. In fact, pollinators were at work millions of years before flowering plants (angiosperms) even existed. In a new review led by Spanish researchers David Peris and Ricardo Pérez-de la Fuente, to which I added a modern ecological perspective, we explored this topic and why it’s relevant to our current understanding of plant-pollinator relationships.
Despite centuries of research on pollination, the fossil record of pollinating insects has only gained serious attention in the past few decades. What palaeontologists have uncovered is reshaping our understanding of pollination’s origins. It turns out that insects were pollinating plants long before flowers evolved—playing a crucial role in the reproduction of ancient gymnosperms, the group of seed-producing plants that includes conifers, cycads, and ginkgos.
Most people assume that insect pollination began with flowering plants, but the evidence tells a different story. Fossilised insects with specialised body structures for carrying pollen—such as hairy bodies or mouthparts adapted for nectar-feeding—have been found in deposits dating back hundreds of millions of years. These early pollinators likely visited gymnosperms, helping them reproduce in a world that looked vastly different from today’s landscapes.
Ancient pollination was driven by a diverse range of insects, many of which are now extinct. The fossil record reveals that various insect groups—including beetles, flies, wasps, and even some long-lost relatives of modern lacewings—were already acting as pollinators long before the first flower bloomed. This means that pollination as an ecological process has far deeper evolutionary roots than many realise.
As plants evolved, so did their pollinators. The rise of flowering plants during the Cretaceous period (around 100 million years ago) transformed pollination systems, leading to the incredible diversity of plant-pollinator relationships we see today. Many of the insect groups that once dominated pollination in prehistoric times have since declined or disappeared, replaced by the bees, butterflies, and other familiar pollinators that thrive in modern ecosystems.
Understanding this long history is essential—not just for scientists, but for anyone interested in biodiversity and conservation. When we focus only on present-day pollinators and plants, we miss a crucial part of the story. The fossil record helps us see how pollination has changed over time, which in turn can offer insights into how today’s ecosystems might respond to environmental pressures such as climate change and habitat loss.
Recognising the ancient history of insect pollination isn’t just an academic exercise—it has real-world implications. If we understand how pollination evolved and adapted to past environmental changes, we can better predict how it might shift in the future. Conservation efforts that aim to protect pollinators today can benefit from a long-term perspective, ensuring that we’re not just responding to recent trends but also considering deep-time ecological processes.
So the next time you see a bee visiting a flower, remember—you’re witnessing the latest chapter in a story that began hundreds of millions of years ago. The relationship between plants and pollinators is far older, more complex, and more fascinating than we ever imagined.
Here’s the reference with a link to the paper. It should be open access, but if you have problems obtaining it, send me a message via my Contact page: