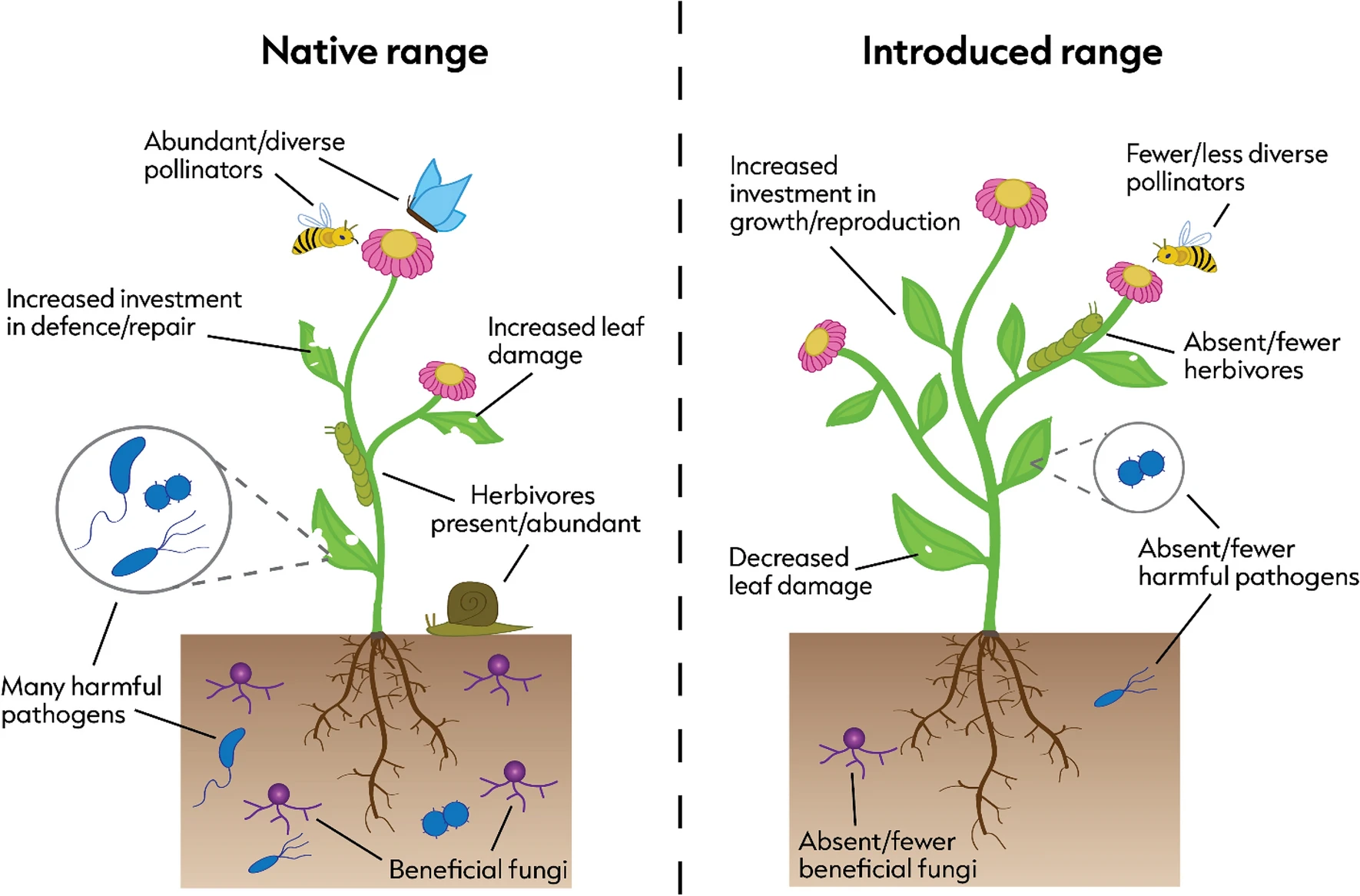
Christmas is a time of expectations, when we assume that familiar traditions will work as they always have. Some traditions are associated with plants such as holly and mistletoe. In ecology, we make similar assumptions, one of the most enduring of which is that plants introduced to new parts of the world often thrive because they leave their natural enemies behind. The thinking goes that leaves that are tough, bitter, or spiky (e.g. holly) or chemically well-defended (e.g. mistletoe), should only strengthen that advantage.
But as with many things we take for granted at this time of year, this intuition does not always hold.
Over the past few years I’ve been part of a large international collaboration led by Zoe Xirocostas (as part of her PhD work) and Angela Moles (see here and here for example) asking deceptively straightforward questions such as: can we predict which introduced plants will experience this “enemy release” based on how well defended their leaves are? In other words, if a plant is tough, hairy, bitter, toxic, or nutritionally poor, does that make it more or less likely to escape herbivores when it turns up somewhere new?
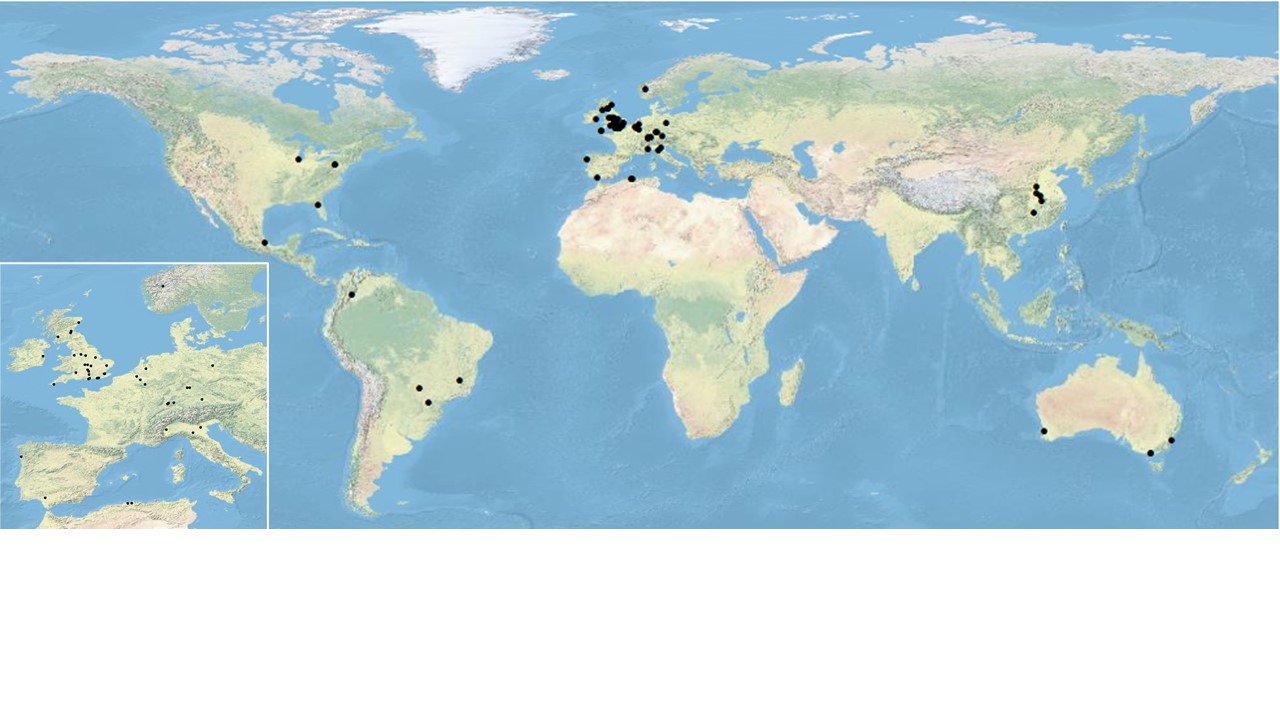
The latest paper from that work has just been published in the journal Oecologia. In it we focused on 16 widespread plant species that are native to Europe but have become established in Australia. For each species, we measured how much leaf damage they suffered in their home range and how much in their introduced range. Our earlier work had already shown that, on average, these plants suffer far less herbivory in Australia – clear evidence of enemy release.
The next step was to ask why.
So we measured an unusually wide range of leaf traits linked to palatability and defence. Some were physical: leaf toughness, thickness, dry matter content, hairiness. Others were chemical: nitrogen content, lipids, phenolics, cyanogenic compounds – the kinds of substances that make leaves bitter, toxic, or difficult to digest. We also combined these into overall “chemical”, “physical”, and “total defence” scores.
If plant defence theory works the way we often assume, at least some of these traits should have explained why certain species escape enemies more than others.
They didn’t.
Across all of these traits – individually and in combination – we found no relationship at all with the strength of enemy release. Plants with heavily defended leaves did not escape herbivores any more than plants with relatively soft, palatable foliage. Equally striking, leaf defences didn’t even predict how much damage plants suffered overall.
This was a genuinely surprising result, especially given how central plant defence traits are to ecological thinking. It suggests that when plants are introduced to new regions, the details of leaf chemistry and toughness may matter far less than we assumed.
One possibility is that context trumps traits. Herbivory may depend more on which insects happen to be present locally, how closely related the newcomer is to native plants, or how chemically novel it is relative to its neighbours, rather than on how well defended it is in absolute terms. Another is that many defensive traits serve multiple roles – coping with drought, heat, or pathogens – and are not just about deterring herbivores. As an example, consider cactus spines, that not only defend a plant against hungry herbivores, but also provide shade for the stem, collect water from fog or dew, and insulate the plant against extreme hot or cold temperatures.
From a practical perspective, our findings have implications for managing invasive species. Biological control programmes – such as the ones that are targeting Japanese Knotweed and Himalayan Balsam in the UK – often assume that poorly defended plants are easier targets. Our results suggest that defence traits alone may not be a reliable guide.
What makes this study particularly powerful is its scale. The same species were measured using the same protocols in different countries; fieldwork was coordinated across multiple climates and ecosystems; statistical analysis drew on shared expertise. This kind of international cooperation can be slow, complex, and occasionally frustrating – but it is exactly what is needed if we want to move beyond neat ideas tested in a single place.
Ecology advances not just through clever hypotheses, but by discovering when our intuitions fail. In this case, a widely held assumption about plant defences and enemy release simply didn’t hold up. That doesn’t close the book on the question – but it does force us to turn the page and rethink what really determines success and failure when species move around the globe.
If you are interested in the full study, here’s the reference with a link to the paper, which is open access:
A Happy Christmas and a Prosperous New Year to all of the readers of my blog!