The region of South America that we know as “Amazonia” has arguably the greatest biological diversity of any part of the planet, certainly as far as plants are concerned. In some places the number of tree species per hectare exceeds 400, an order of magnitude greater than the number for the whole of the British Isles. However estimating the total number of even the described plant species in this vast area has proven controversial, as two recent studies exemplify. The first study was by ter Steege et al. (2016) and entitled “The discovery of the Amazonian tree flora with an updated checklist of all known tree taxa“, whilst the second is from just last month: Cardoso et al. (2017) “Amazon plant diversity revealed by a taxonomically verified species list“. Both of them are open access so click on the links if you want to read the full studies.
One might expect that two such studies focused on Amazonia, both using vouchered herbarium records, would reach broadly similar conclusions as to the number of tree species in the region. Not a bit of it: ter Steege et al. (2016) report 11,676 species, whilst Cardoso et al. (2017) say that the figure is 6,727. That’s almost a two-fold difference! Why the discrepancy? Inspired by an initial tweet by University of Glasgow taxonomist Roderic Page, I downloaded the data from both studies and looked at it closely.
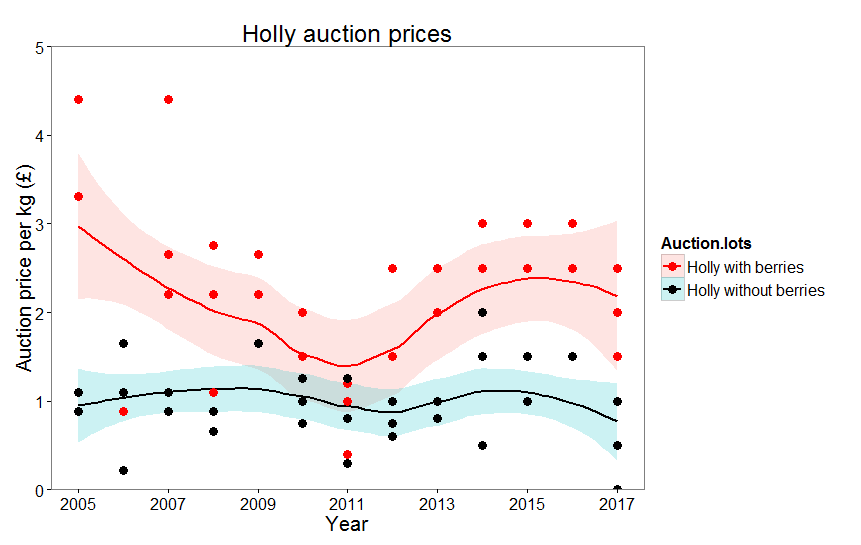
Here’s a scatter plot of the number of tree species per plant family reported by both studies:
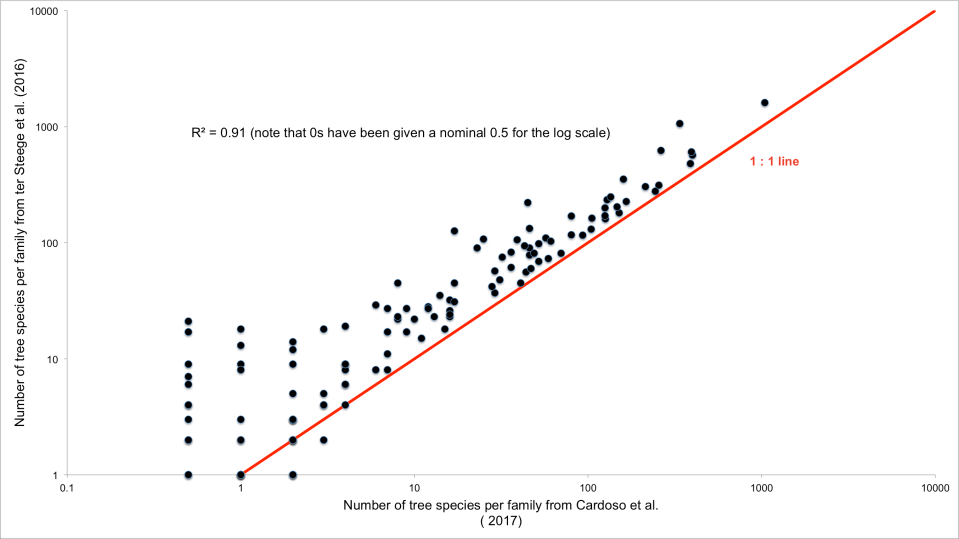
The red line shows where we would expect the data points to lie if both studies had reported the same number of tree species per family. Clearly few families lie on this line and most are above it as we might expect: as I’ve said, ter Steege et al. (2016) concluded that there were far more tree species overall and this is reflected at the family level. Note that I’ve graphed this using a log scale and what might seem to be small differences are actually very large indeed.
Although the findings from two studies are highly correlated (diverse families are diverse in both studies, ditto families with low diversity) the actual level of that species richness is very different. For example, in the Annonaceae, ter Steege et al. report 480 species, Cardoso et al. report 388; in the Clusiaceae the figures are 247 versus 135. Other families are excluded from one data set or the other: ter Steege et al. reckon there 7 species of trees in the Dilleniaceae whereas Cardoso et al. cite zero. Here’s a link to the data set if you want to explore further.
So what’s going on here? Why do two studies with similar aims, published about 12 months apart, come to such different conclusions. As far as I can see there are three reasons for this.
First of all, the studies used slightly different taxonomies when it came to considering families and species. So for example, Cardoso et al. recognise the family Peraceae which ter Steege et al. do not. Although I haven’t done it, I’m sure that if one were to dig down to the species level there would be differences in which species were accepted and which were considered synonyms.
Secondly, the exact definition of what constitutes a “tree” varies between botanists, and the non-botanists who are no doubt responsible for some of the plant collections: some consider anything to be woody and tall-ish to be a “tree”, others have more strict definitions. Notes about growth form taken in the field consequently get included in herbarium databases and may be inaccurate, especially for the uncommon species that have rarely been seen in the field.
The final reason, and the one that seems to be responsible for most of the discrepancy, is the definition of what constitutes “Amazonia”. In the first study ter Steege et al. defined it as including the “forests and savannahs of the Amazon basin and Guiana Shield”. In contrast Cardoso et al. considered only “lowland Amazon rain forests”. That’s a big difference as there’s lot of savannah in this region, as well as other habitat types. When we did field work in Guyana some years ago we could travel very quickly between savannah and rainforest. It was clear to us that there is a range of trees that are restricted to one habitat or another, including species of Dilleniaceae (mentioned above) that are savannah specialists (hence the family’s exclusion from the Cardoso et al. study).
Now neither of these studies is “wrong” in the sense of being inaccurate or misguided: both are great studies involving a huge effort on the part of the authors. But the limitations and definitions of geography and taxonomy that I’ve highlighted do mean that they need to be treated as rather different and not directly comparable.
So how many tree species are there in Amazonia? If we consider just the rainforest then it’s 6,727 (Cardoso et al. 2017). If we consider all habitats in the region, including rainforest plus savannah etc., then the figure is 11,676 species (ter Steege et al. 2016). One of the implications of this is that the non-rainforest “Amazonian” habitats collectively contain 4949 tree species. Thus a large proportion of the diversity of the region is in habitats, such as savannah, which are less of a focus for conservation efforts and not as well known to the general public, but are at least as threatened by agriculture and mining as rainforest.
Thanks to Roderic Page for initially highlighting this on Twitter, and Sandy Knapp for discussion.
UPDATE: In retrospect my conclusion above regarding the proportion of trees in non-lowland rainforest habitats was much too high, as a couple of commenters have noted below. It’s worth reading what they have to say, and my responses. It’s likely that the taxonomic differences between the two studies are at least as great as the geographical ones, but then taxonomic opinions vary hugely. Just serves to emphasise what a controversial and problematic question this is!