When we think of cities, gardens might not be the first thing that comes to mind. But these green patches — whether in private yards, parks, or balconies — play a surprisingly important role in supporting urban wildlife. Among their most crucial guests? Pollinators like bees, butterflies, and even birds and bats.
In a new study just published, I teamed up with some Brazilian colleagues to explore how the different features of garden flowers help sustain pollinators throughout the year in a subtropical urban garden. While we’ve long known that garden flowers provide food for pollinators, what’s less clear is how specific floral traits — like shape, flowering time, and type of nectar or pollen — influence who visits which plants and when.
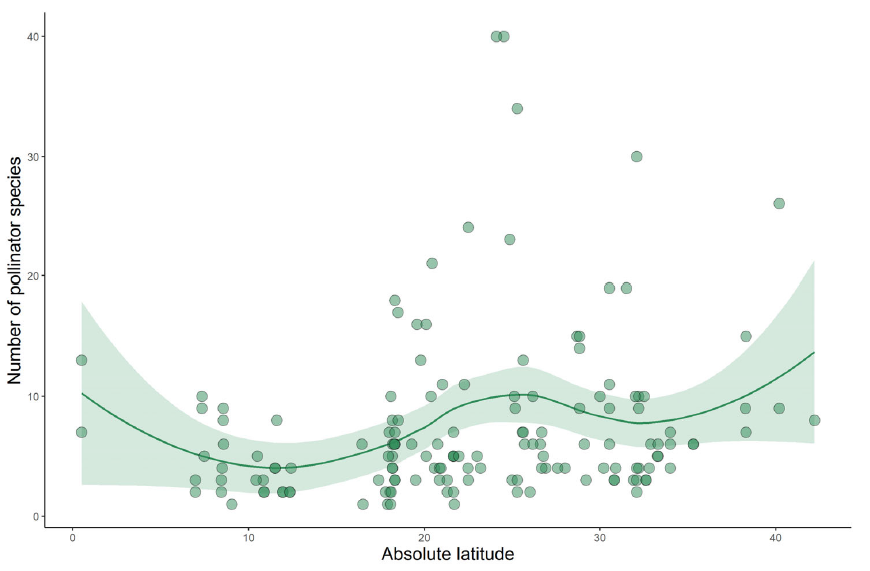
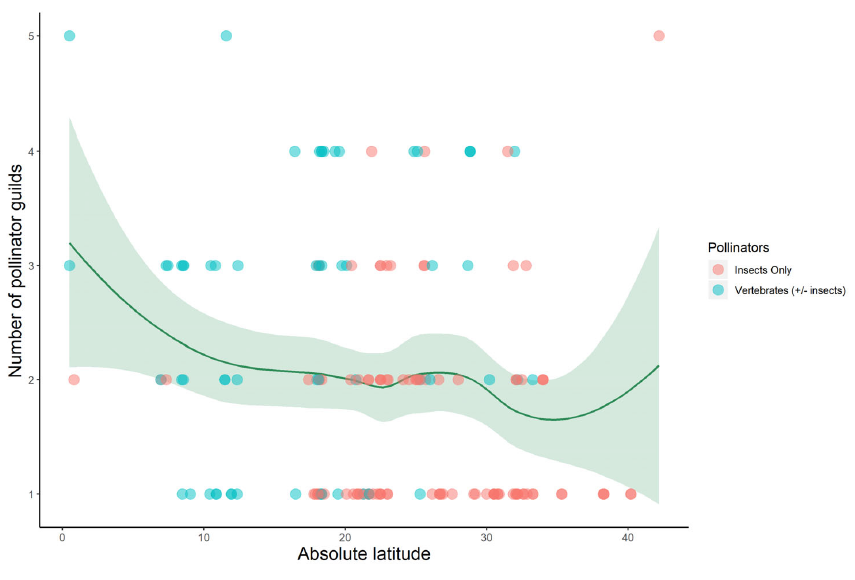
To get a clearer picture, we conducted weekly surveys of pollinators visiting garden flowers over the course of a year. We paid close attention to traits such as the depth of flower, the kind of resources offered (nectar vs. pollen), how closely related different plants were, and when they flowered.
What we found was striking: the network of interactions between flowers and pollinators was highly organized. Plants grouped into clusters, or “modules,” that tended to share similar physical traits and evolutionary histories — but interestingly, not the same flowering times. This meant that within each module, different plants flowered at different times of year, effectively staggering their blooms so that there was always something on offer for pollinators.
Even more intriguing was the discovery that most plants had just a few connections in the network, usually restricted to a single module. These “peripheral” plants accounted for over 85% of all pollinator visits. Meanwhile, a few special species acted as bridges between modules — their role in linking different parts of the network made them key to its stability. These connector species didn’t flower at the same time, which helped to maintain a steady supply of food for pollinators across seasons.
Not all interactions between plants and pollinators are “legitimate” in the sense of leading to pollination. Some animals visit flowers just for the food, without helping with reproduction. But our study found that these interactions still played a valuable role in supporting a diverse pollinator community.
So what does all this mean for urban gardeners and city planners?
First, it highlights how important it is to plant a variety of flowers that bloom at different times of year. Second, it shows that even seemingly minor plants or interactions can contribute to the ecological resilience of urban green spaces. And finally, it underscores that thoughtful planting — considering things like flower shape, blooming schedules, and diversity — can help keep pollinators thriving, even in the heart of the city.
Urban gardens aren’t just pretty — they’re powerful allies in the fight to support biodiversity.
The study was led by Brazilian research student Luis de Sousa Perugini. Here’s the reference with a link to the paper:
Here’s the abstract:
Garden flowers play a vital role in urban environments, supporting pollinator communities. Yet, the extent to which floral traits shape urban pollination networks remains poorly understood. This study investigated how garden plants shape year-round pollination networks, sampled in weekly surveys in an urban subtropical garden. We focused on the role of floral morphology (corolla depth), type of resource, relatedness, and phenology in the organization of interactions. We determined whether modularity and species roles were influenced by these floral traits, comparing if legitimate pollination, illegitimate (i.e. non-pollinating) interactions and all interactions had similar drivers. All networks were modular, and in the overall network plants within the same module were morphologically and phylogenetically similar while their phenology was significantly overdispersed throughout the year. Peripheral species, those with few interactions and restricted to a single module, dominated all networks, representing over 85% of interactions. We found that phenology was related to the species role of overall network connectors (species that connect modules) and legitimate module hubs (species that connect their own modules). Both showed no overlap in their flowering periods, providing floral resources at different times of the year. Each module functioned as a distinct unit, showing year-round availability of resources to support its pollinators. This suggests that resource continuity and trait-based filtering may shape pollinator assemblages influencing ecological resilience in urban habitats. Even interactions that do not contribute to plant reproduction can sustain a diverse fauna, highlighting the importance of these interactions in urban green space planning and management.