One of the projects in which I’m currently involved is the WorldFAIR project. Funded by the European Commission, WorldFAIR is exploring how to make data FAIR – Findable, Accessible, Interoperable and Reusable – across a range of different disciplines in the sciences and humanities.
My involvement is specifically with Work Package 10, which is focused on data standards for plant-pollinator interactions, particularly as they relate to pollination of agricultural crops. After a year of hard work, I’m delighted to announce that our interim draft report from this Work Package has just been published! You can read the summary and download the report from Zenodo – here’s the link: https://zenodo.org/record/8176978
There’s more to come over the next twelve months and I’ll post updates as and when they appear. In the meantime, do check out the WorldFAIR website for information about the other Work Packages, their webinar series, FAIR data standards, and so forth.
Earlier this week, the East Midlands Environment Agency proudly tweeted that they had placed honey bee hives on an ecologically important site that they own. As you might imagine, the response from pollinator experts such as myself, conservation NGOs, and some beekeepers, was not positive, as you can see if you look at the comments beneath my tweet:
If this is an important site for nature conservation, as claimed, then the #EnvironmentAgency should NOT put hives of honey #bees on it!
— Prof. Jeff Ollerton – @JeffOllerton@ecoevo.social (@JeffOllerton) July 12, 2023
By coincidence, overnight I received a message from someone in the USA asking for advice. Here’s a redacted version of their message:
My community has a 4 acre serpentine barren site that is part of a larger string of these unique barrens ….. Honey bee hives have recently been located adjacent to the barrens. Can you advise me as to the best way to determine whether there are, and to document any, adverse effects to the serpentine barrens native pollinators?
Going back to the question of how to assess any impacts, the simple answer is that it’s not easy and it relies on having good data. This was my response to my American correspondent:
Ideally you would need to take a before-and-after approach where you have data on things like number of native pollinator species, their abundance (including nest sites), rates of visitation of different pollinators to flowers, and fruit or seed set from particular plants. You’d then compare what was going on before the hives arrived with what’s occurring since their arrival.
If you don’t have the “before” data it’s much more difficult to assess if there has been an impact from the honey bees. However, the advice of most conservation groups is to adopt the “precautionary principle” and not site hives on or adjacent to areas of nature conservation value, especially if they are relatively small areas. See for example the Bumblebee Conservation Trust’s advice: https://www.bumblebeeconservation.org/managed-honeybees/
The precautionary principle is a well established concept across a range of areas, including health and engineering, as well as nature conservation. In the latter it needs to be more widely applied, especially when it comes to questions of where to site honey bee hives, and how many.
At the moment Karin and I are in the UK for a couple of weeks. I had work to do as an external examiner at the University of Swansea, plus we wanted to catch up with some family and friends. Our main base has been the home of our mates Ian and Simone and we’ve enjoyed some warm, muggy evenings sitting in their garden chewing the fat. Every now and again my eyes have been drawn to the activities of bumblebees as they move in and out of the foliage of a small Silver Fir. The bees are attracted to the large colonies of an aphid that is feeding on the tree’s trunk, from which they are collecting honeydew, as you can see in the photograph above.
When we think of the diets of bees we automatically think “nectar and pollen”. Honeydew, as a sugar-rich fluid, fits broadly into this concept, though as far as I know there’s been little study of its relative importance as a food source for bees. Aside from a few “vulture bees”, all of the 20,000 or so species are vegetarian. And therein lies a problem. Bees evolved from carnivorous wasps and so the evolution of bees, and their complex ecologies, is tied into this profound dietary shift toward a plant-based diet.
A particular issue that has hardly been investigated until recently is that the ratio of elements within meat is very different to that of plants. In particular, animal tissue has a high ratio of sodium (Na) relative to potassium (K), whereas for plants the ratio is reversed – high ratio of K:Na.
In a new conceptual review paper with my colleagues Zuzanna Filipiak and Michał Filipiak, we have explored the implications of this difference in elemental ratios for bee ecology and evolution, and for the conservation of these important insects. The paper is open access and you can download a copy by following a link in this reference:
Bees provide important ecological services, and many species are threatened globally, yet our knowledge of wild bee ecology and evolution is limited. While evolving from carnivorous ancestors, bees had to develop strategies for coping with limitations imposed on them by a plant-based diet, with nectar providing energy and essential amino acids and pollen as an extraordinary, protein- and lipid-rich food nutritionally similar to animal tissues. Both nectar and pollen display one characteristic common to plants, a high ratio of potassium to sodium (K:Na), potentially leading to bee underdevelopment, health problems, and death. We discuss why and how the ratio of K:Na contributes to bee ecology and evolution and how considering this factor in future studies will provide new knowledge, more accurately depicting the relationship of bees with their environments. Such knowledge is essential for understanding how plants and bees function and interact and is needed to effectively protect wild bees.
But just as when a movie director says “That’s a wrap” at the end of the final day of filming, the hard work does not stop here. Two people have read the full manuscript as I was producing chapters and their suggestions have been incorporated into this draft. The publisher will now send it to a third, independent beta reader and once their feedback has been acted on it will go to a copy editor who will suggest stylistic changes, check for logic and consistency, and so forth.
At the same time I will be choosing which plates to put in the book, which images to use on the back cover, writing their descriptions and deciding where to cite them; checking the sources and further reading sections for each chapter and formatting the references; and producing an appendix that lists the scientific names against the vernacular names that I am using in the book. I also need to finalise the acknowledgements section.
As an author, producing a book is a long process that doesn’t end with the actual writing of the manuscript. It’s incredibly satisfying, however, and working with Pelagic on my second book for them has been a great experience. All being well, Birds & Flowers should be out by early winter.
Now, I have three options for the next book that I’m writing….which one to choose…?
As a teenager one of my main interests was collecting fossils. In search of specimens I wandered for hours, scouring the Carboniferous coal shale heaps and Permian reef outcrops of my native Sunderland. I spent so much time bothering the geology curator at the local museum with my inquiries that he offered to host me for a year as the placement part of my college course. If I had been able to convince my tutors that paleontology was really just biology in deep time I may have ended up as a professional fossil researcher. But it was not to be and instead I spent a (mostly happy) year working in the microbiology laboratory of a local brewery.
My interest in the ecology of the past has never left me, and over the years I’ve contributed a few articles to journals commenting on the latest fossil findings as they relate to pollination and flowering plant evolution. So I was delighted to be asked by Spanish paleontologist David Peris to help with a new review of insect pollination in deep time, led by PhD candidate Constanza Peña-Kairath. That review has just been published in Trends in Ecology & Evolution, and for the next 50 days it’s available for free download by following the link in the reference:
Inferring insect pollination from compression fossils and amber inclusions is difficult because of a lack of consensus on defining an insect pollinator and the challenge of recognizing this ecological relationship in deep time. We propose a conceptual definition for such insects and an operational classification into pollinator or presumed pollinator. Using this approach, we identified 15 insect families that include fossil pollinators and show that pollination relationships have existed since at least the Upper Jurassic (~163 Ma). Insects prior to this can only be classified as presumed pollinators. This gives a more nuanced insight into the origin and evolution of an ecological relationship that is vital to the establishment, composition and conservation of modern terrestrial ecosystems.
This is a guest post by Dr Annemarie Heiduk about a new species that she’s recently described.
—————————————————————————————————————————-
In an earlier blog post about the discovery of a novel Ceropegia species, Jeff’s finishing sentence was: “I wonder what else is still waiting to be discovered in the stunning grasslands of South Africa?”
I am happy to provide a first answer to this question: Ceropegia stylesii.
This new species does not have the tubular kettle-trap flowers typical of Ceropegia, which temporarily trap pollinators, but open-rotate corollas where the gynostegium (fused male and female reproductive organs) is freely accessible to pollinators. So, in the traditional sense, C. stylesii is not a Ceropegia. This needs some explanation here!
Ceropegia is a genus in the plant family Apocynaceae (Jeff’s favourite family of plants!) and therein the genus is placed in the subfamily Asclepiadoideae which originally was a family on its own (“Asclepiadaceae”).
Within this subfamily, the genus Ceropegia belongs to the Stapeliinae – a subtribe which comprises ca. 720 species. About 220 species thereof have exciting looking and very cleverly designed kettle-trap flowers which attract small flies as pollinators via deceptive strategies (see http://plantlifesouthafrica.blogspot.com/2019/07/plantlife-sa-volume-473-july-2019.html). The remaining species in Stapeliinae are the well-known stem-succulent stapeliads (ca. 355 species in >30 genera) and ca. 140 species known as Brachystelma.
With increasingly better molecular methods to study the evolutionary relationships of species in Stapeliinae, the traditional grouping of the species was illuminated as being artificial, i.e., species with kettle-trap flowers are not actually a natural group and Brachystelma species are scattered among them; the stapeliads are also nested in Ceropegia but as a single (monophyletic) group. These results based on DNA-sequence similarities are not compatible with the traditional generic concept in Stapeliinae, and as a result, changes were instigated.
Some colleagues wish to see all 720 species of Ceropegia, Brachystelma and the stapeliads merged into one single large genus Ceropegia, a solution which would entail more than 400 new name combinations. Others prefer to adopt a less dramatic change of concept and only include Brachystelma in an enlarged Ceropegia while keeping the stapeliads separate based on their monophyly and distinct vegetative features. This pragmatic solution considers both taxonomic and phylogenetic facts and reduces the previously multiple cases of paraphyly to a single case. More importantly, it avoids hundreds of name changes in the group. Both concepts are correct in their own right and justified, so it is a personal decision which one to follow.
The newly described species C. stylesii would traditionally have been placed in Brachystelma as it is lacking tubular kettle-trap flowers. After the inclusion of Brachystelma into Ceropegia, C. stylesii is placed within section Bowkerianae – a group comprising species both with and without tubular kettle-trap flowers. With the description of C. stylesii, the section now has 15 members of which 10 have open-rotate flowers. Among these, C. stylesii appears to be most closely related to C. gerrardii from which it can only readily be distinguished when in flower (see the lower most image above).
The flowers of C. stylesii superficially look like miniature versions of a dark-flowered form of C. gerrardii, which growths in the same habitat. C. stylesii flowers are only about 6 mm in size whereas those of C. gerrardii are about three times larger. While C. gerrardii occurs in grasslands throughout eastern South Africa, C. stylesii is believed to be endemic to Ngome, where it is known from two localities with a total of less than 10 plants. After the recent discovery of C. heidukiae at Ngome, the area revealed another outstanding member of this amazing plant group, and thereby once again proves its conservation importance.
C. stylesii is named for David Gordon Alexander Styles, botanical explorer and collector, to honour his valuable contribution to botanical knowledge in South Africa. David is renowned for “…his daring nature to go leaps and bounds for the specimen he is interested in” (see Chetty 2021), a statement I can readily confirm based on personal experience. Many of David’s collections (by now well over 6000 specimens donated to various herbaria) are novelties awaiting to be described. With C. stylesii, a total of five plant species bear his name. I am delighted that eventually a Ceropegia species could be named for him as David’s knowledge on the distribution and habitats of these special plants is of great value to my research on this plant group.
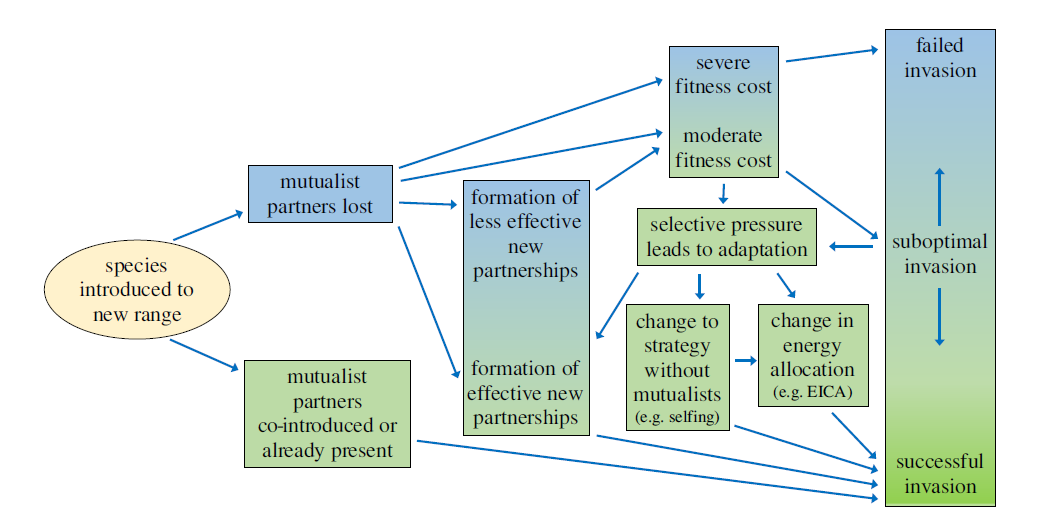
All organisms – be they plants, animals, fungi, or whatever – interact with other species throughout their lives, in relationships that include predation, parasitism, commensalism, and the many and varied forms of mutualism. But when species are transported to a different part of the world, as has happened often during the Anthropocene, these interactions typically break down because usually only one of the participants moves. This loss of ecological relationships can play a role in whether or not a species becomes established in its new home, and has been mostly explored in the “Enemy Release Hypothesis” (ERH) which predicts that, by leaving behind predators or parasites or herbivores, a species becomes more ecologically successful and ultimately invasive in its novel range.
Less well studied, though potentially just as important, is the “Missed Mutualist Hypothesis” (MMH) which in a sense is the twin of the ERH. As well as leaving behind “enemies”, introduced species leave behind “friends” such as pollinators, seed dispersers, mycorrhizal fungi, defensive partners, and other mutually beneficial associates. Negative effects arising from the loss of these relationships could potentially balance the positive impacts arising from the ERH.
In a new quantitative review just published, we review what’s known about the MMH (currently much less than the ERH) and suggest some fruitful lines of enquiry. The study is led by Angela Moles, my collaborator at the University of New South Wales where I spent time as a Visiting Research Fellow in 2019/20 (see my blog posts about that visit starting here). The paper has had a long gestation and gone through several iterations and revisions since we started writing it in late 2019, not least caused by the covid pandemic, but I think that it’s all the better for it.
Here’s the full reference with a link to the paper:
Introduced species often benefit from escaping their enemies when they are transported to a new range, an idea commonly expressed as the enemy release hypothesis. However, species might shed mutualists as well as enemies when they colonize a new range. Loss of mutualists might reduce the success of introduced populations, or even cause failure to establish. We provide the first quantitative synthesis testing this natural but often overlooked parallel of the enemy release hypothesis, which is known as the missed mutualist hypothesis.
Meta-analysis showed that plants interact with 1.9 times more mutualist species, and have 2.3 times more interactions with mutualists per unit time in their native range than in their introduced range. Species may mitigate the negative effects of missed mutualists. For instance, selection arising from missed mutualists could cause introduced species to evolve either to facilitate interactions with a new suite of species or to exist without mutualisms. Just as enemy release can allow introduced populations to redirect energy from defence to growth, potentially evolving increased competitive ability, species that shift to strategies without mutualists may be able to reallocate energy from mutualism toward increased competitive ability or seed production. The missed mutualist hypothesis advances understanding of the selective forces and filters that act on plant species in the early stages of introduction and establishment and thus could inform the management of introduced species.
The question of whether interactions between different species are more specialised in tropical environments (as theory predicts) has intrigued me for a couple of decades. In fact it’s just occurred to me that August 2022 was the 20th anniversary of my paper in Oikos co-authored with Louise Cranmer entitled: Latitudinal trends in plant-pollinator interactions: are tropical plants more specialised? That paper was one of the first to seriously challenge an idea that was long-embedded in the scientific and (especially) popular literature, that tropical ecology was in a sense “special” and that the ways in which species parasitised, consumed, or engaged in mutualistic relationships in the tropics was different to what was happening in the subtropics and temperate zones.
Since then I’ve written about this subject in a number of publications, most recently in my book Pollinators & Pollination: Nature and Society and it’s inspired some other researchers to address the topic.
One of the real challenges with asking questions about how plant-pollinator relationships change over large geographical areas is obtaining good, robust data to analyse. It’s a challenge to convince science funding agencies to give money to spend many years travelling the world and collecting the kind of data that are needed. However we can gain some idea of the patterns, and potential processes, that drive the macroecology of plant-pollinator interactions by piecing together databases of interactions for particular taxa, gleaned from published and unpublished sources.
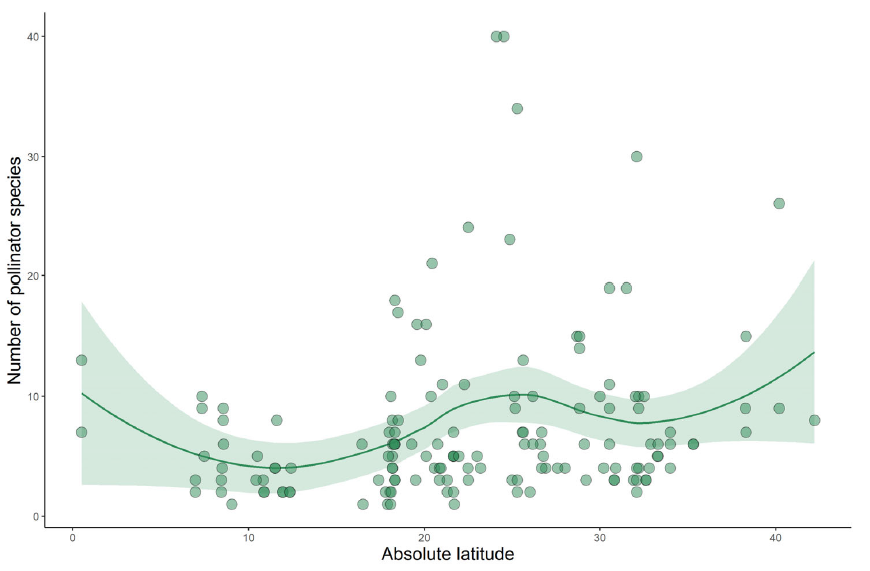
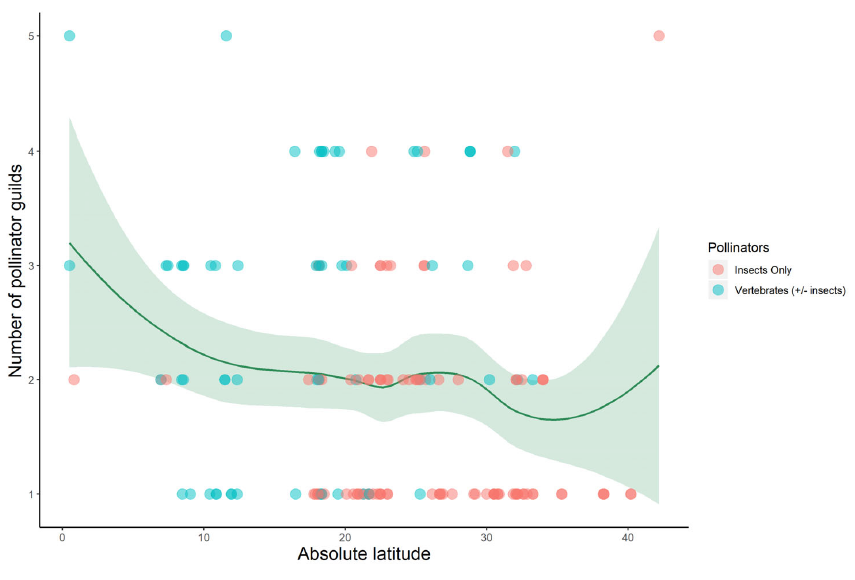
That’s what we have done for the family Cactaceae in a new study led by Pablo Gorostiague from the Universidad Nacional de Salta in Argentina. This collaboration started when Pablo visited Northampton back in 2018 and spent some time with my research group, including helping out with field work in Tenerife. Since then the usual issues (work, COVID, etc.) have delayed publication of our paper, but now it’s finally out. Amongst other results we find that, yes, tropical cacti are pollinated by fewer species on average (though it’s hugely variable – see the figure above) but that functional specialisation (i.e. the number of pollinator guilds that are used by species) is no different in the tropics compared to the extra-tropics (that’s the figure at the end of this post).
The full reference with a link to the paper is below; if anyone wants a PDF, please send me a message via the Contact page:
Biotic interactions are said to be more specialized in the tropics, and this was also proposed for the pollination systems of columnar cacti from North America. However, this has not yet been tested for a wider set of cactus species. Here, we use the available information about pollination in the Cactaceae to explore the geographic patterns of this mutualistic interaction, and test if there is a latitudinal gradient in its degree of specialization.
We performed a bibliographic search of all publications on the pollination of cacti species and summarized the information to build a database. We used generalized linear models to evaluate if the degree of specialization in cacti pollination systems is affected by latitude, using two different measures: the number of pollinator guilds (functional specialization) and the number of pollinator species (ecological specialization).
Our database contained information about the pollination of 148 species. The most frequent pollinator guilds were bees, birds, moths and bats. There was no apparent effect of latitude on the number of guilds that pollinate a cactus species. However, latitude had a small but significant effect on the number of pollinator species that service a given cactus species.
Bees are found as pollinators of most cactus species, along a wide latitudinal gradient. Bat and bird pollination is more common in the tropics than in the extra-tropics. The available information suggests that cacti pollination systems are slightly more ecologically specialized in the tropics, but it does not support any trend with regard to functional specialization.
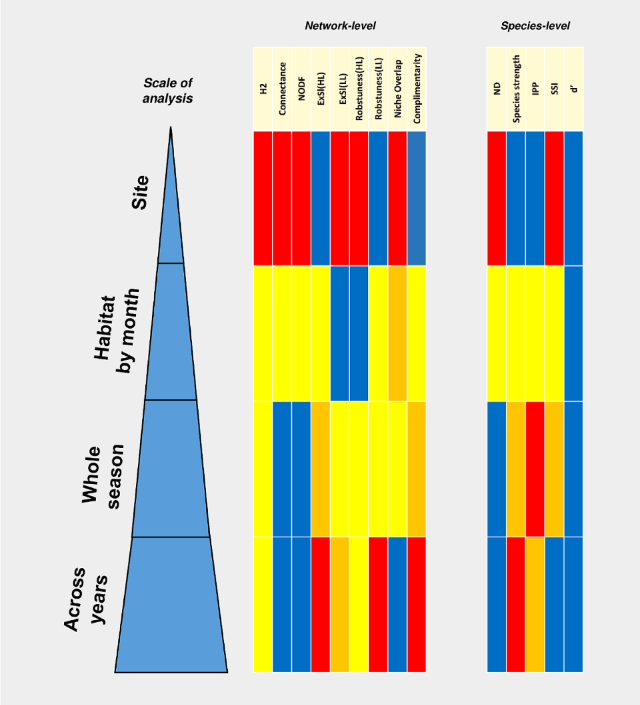
It’s long been recognised that the scale at which we study the natural world – over long or short time periods, or across small areas or whole regions – affects the conclusions that we draw about ecological patterns and processes. This is certainly true of plant-pollinator interactions. For example, a widely distributed plant can have very different pollinators at the extremes of its range, and pollinators like bees may vary their focus on nectar and pollen sources from year to year.
The analysis of these interactions as networks of actors has become increasingly popular in the last couple of decades. However there is no consensus about how frequent sampling should be, or the geographical scale over which networks should be studied. In fact all scales (from regional “meta-networks” down to single-season, single-site, single taxon observations) are relevant, depending on the questions being asked or the hypotheses posed.
But it’s important that we acknowledge that conclusions drawn at one scale may not apply at other scales.
That’s the take home message from a paper published this week which is the latest output from the PhD work of Australian bee expert Kit Prendergast. We have collaborated on several papers based on her data and this is actually my 100th peer-reviewed publication: a proud milestone for me and one which I’m glad to share with a wonderful early career researcher like Kit!
Here’s the reference with a link to a read-only version of the paper:
Bipartite networks of flowering plants and their visitors (potential pollinators) are increasingly being used in studies of the structure and function of these ecological interactions. Whilst they hold much promise in understanding the ecology of plant– pollinator networks and how this may be altered by environmental perturbations, like land-use change and invasive species, there is no consensus about the scale at which such networks should be constructed and analysed. Ecologists, however, have emphasised that many processes are scale dependent. Here, we compare network- and species-level properties of ecological networks analysed at the level of a site, pooling across sites within a given habitat for each month of surveys, and pooling across all sites and months to create a single network per habitat type. We additionally considered how these three scales of resolution influenced conclusions regarding differences between networks according to two contrasting habitat types (urban bushland remnants and residential gardens) and the influence of honey bee abundance on network properties. We found that most network properties varied markedly depending on the scale of analysis, as did the significance, or lack thereof, of habitat type and honey bee abundance on network properties. We caution against pooling across sites and months as this can create unrealistic links, invalidating conclusions on network structure. In conclusion, consideration of scale of analysis is also important when conducting and interpreting plant–pollinator networks.
Within the last decade there’s been a growing awareness of the importance of urban environments for supporting populations of pollinators, especially bees. Indeed, I devoted a whole chapter of my book Pollinators & Pollination: Nature and Society to the topic, though even then I was only able to scratch the surface of the research that’s been done. Since then there’s been some important studies published and this 2020 review by Kath Baldock provides a good starting point for the topic, whilst a recent pre-print by Pietro Maruyama and colleagues emphasises how little we know about pollinators in tropical cities.
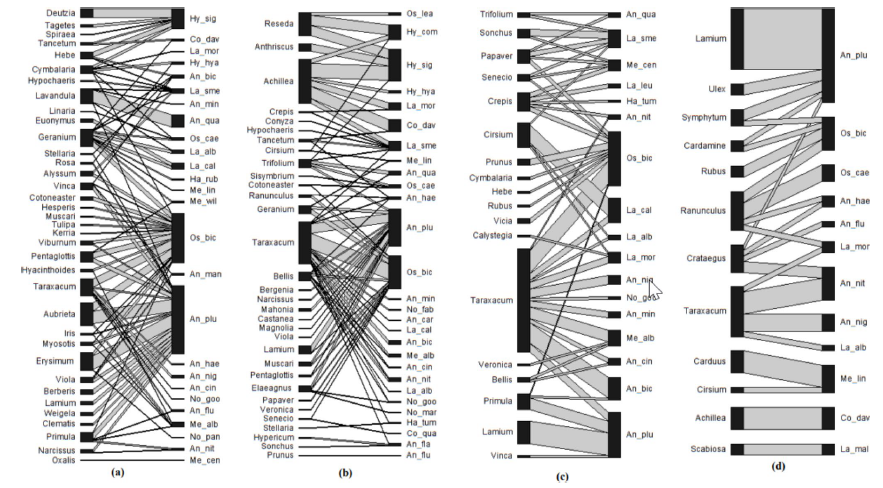
One of the most detailed studies of urban solitary bees in a British town was conducted by Muzafar Sirohi when he was a PhD researcher in my department in Northampton. The first paper from that work, documenting the diversity and abundance of bees, came out in 2015, but since then commitments to other projects, plus Muzafar’s return to his university in Pakistan, have meant that we’ve struggled to find the time to publish more. Hopefully that’s changing and the second publication from Muzafar’s thesis is now out, with a third in progress.
This new paper uses a network approach to study the use of flowers by these bees; here’s the reference with a link to a read-only copy of the paper, followed by the abstract.
Biodiversity is declining through human activities and urbanisation is often seen as a particular concern. Urban settings, however, provide diverse microclimatic conditions for plants and pollinating insects, and therefore may be significant habitats for the conservation of solitary and primitively eusocial bees, a major group of pollinators. This study analysed the interactions between these bees and the plants on which they forage, using a network approach. We compared urban habitats (gardens, roadsides, and open vegetation) in a large British town with nearby nature reserves. One native plant Taraxacum officinale (dandelion) was a core generalist species visited in all habitat types. Other core plant species restricted to particular habitats include species of Geranium, Bellis, Crepis, and Ranunculus. Two generalist bee species, Anthophora plumipes and Osmia bicornis were the core visitor species within the networks. The networks were comparatively more nested in urban habitat types than nature areas, suggesting more frequent interactions between generalist and specialist species in urban areas. Network connectance, network level specialisation (H2’ index), and plant generality (network level) were not significantly different in urban and nature areas. However, visitor generality was found to be significantly higher in urban gardens than in nature areas. Careful management of common urban vegetation would be beneficial for supporting urban wild pollinators.