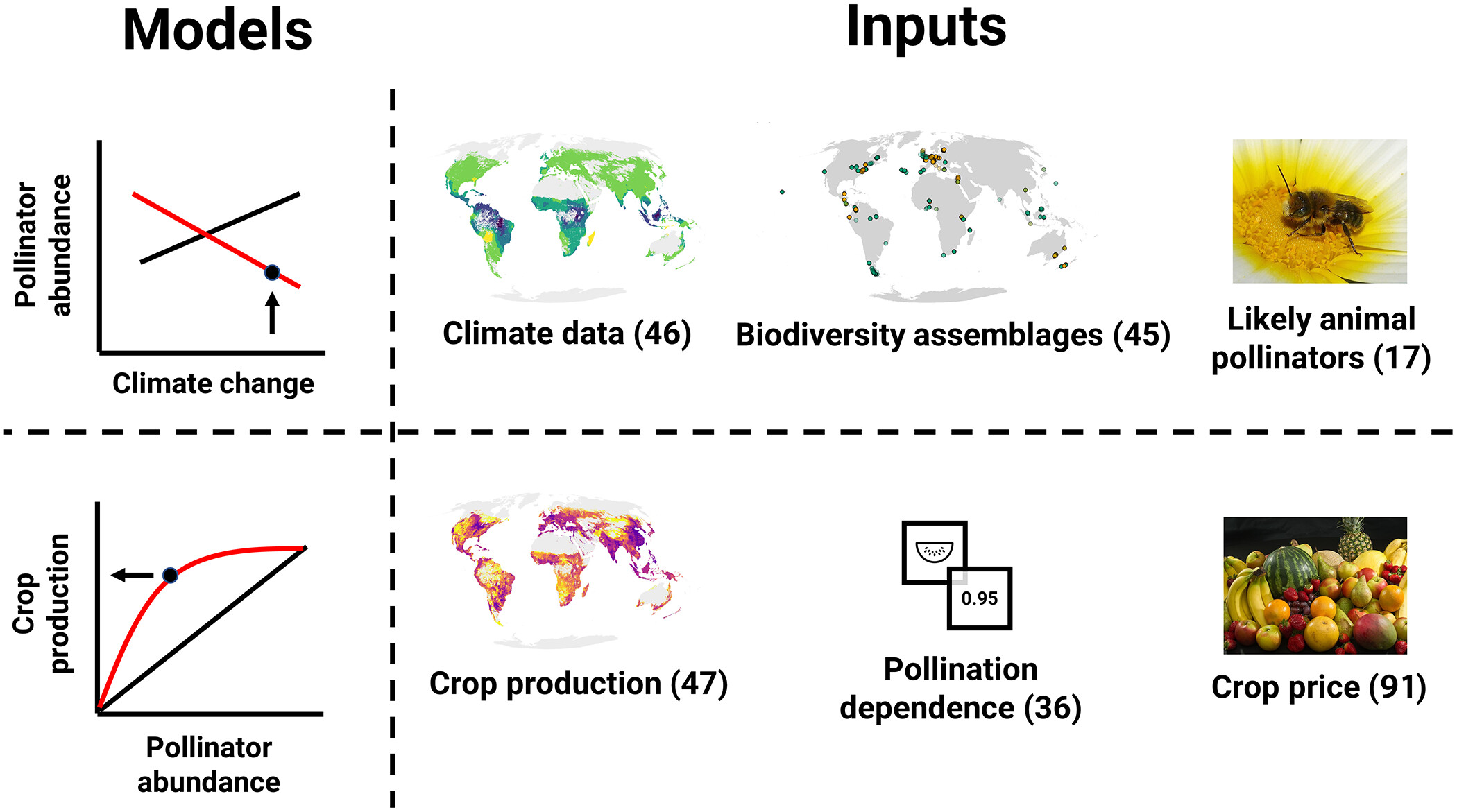
Yesterday I received an email containing the following press release. I thought it might interest readers of the blog so I am copying it in full with no edits. I had a quick look over the report and it’s amazingly detailed and comprehensive. It’s a shame that the report only covers insects, but that probably reflects my current bias given that my next book, due out in February, is about pollinating birds! Press release follows:
Governor Polis and the Department of Natural Resources Release Pollinator Report
BROOMFIELD – Today, Governor Polis in partnership with The Colorado Department of Natural Resources (DNR), Colorado State University Extension, Xerces Society for Invertebrate Conservation, and University of Colorado Museum of Natural History released the Colorado Native Pollinating Insects Health Study which is the most robust and detailed account of pollinator health ever undertaken in Colorado history. As directed by SB22-199, Native Pollinating Insects Protection Study, sponsored by Senators Sonya Jaquez Lewis and Kevin Priola, and Representatives Cathy Kipp and Meg Froelich, signed by Governor Polis on May 27, 2022, the study assesses the health of Colorado’s native pollinators, evaluates state policies for safeguarding pollinators, and makes recommendations on how to preserve and protect pollinators in Colorado.
“Pollinators play a critical role in Colorado life. From Crested Butte’s beautiful spring meadows to Palisade Peaches and Rocky Ford melons, Colorado’s pollinators sustain our flora and enable many foundational industries in every corner of the state. As our climate changes, we must safeguard the pollinators that generate and regenerate the Colorful Colorado we love,” said Governor Polis
Colorado is home to various native insects and bats whose pollinating services are at the heart of healthy environments and economies. Pollinators are critical to Colorado’s economy and our agricultural production and food systems, and they are essential for flowering plants that support the state’s wildlife ecosystem and add color to Colorado’s beautiful landscapes.
“Colorado is fortunate to have a tremendous diversity of plants and animals, but pollinating insects are perhaps the least studied but most beneficial for our ecosystems, economy and quality of life,” said Dan Gibbs, Executive Director, Colorado Department of Natural Resources. “I greatly appreciate the time and effort of the study authors who truly did a deep dive into the current state of pollinating insects and state policies and structures. I look forward to working with Colorado legislators and stakeholders in pursuing the best policies to ensure pollinating insect protection and long-term health.”
Colorado is home to over 1,000 species of bees—nearly 30% of North America’s and approximately 5% of the world’s bee species—and nearly 300 species of butterflies, representing over 40% of the diversity of butterflies in North America north of Mexico, some of which are already listed under the federal Endangered Species Act. This report underscores the importance of the Polis administration’s goals to tackle Colorado’s greenhouse gas emissions, prepare industries for the impending effects of climate change, and create more sustainable living in Colorado for pollinator populations.
“Our ecosystems rely on pollinators, which is why I’ve championed measures that limit toxic chemicals from harming pollinators and put forth a statewide assessment to better understand the problems our pollinators face and identify practices to better sustain them,” said Senator Sonya Jaquez Lewis, D-Longmont. “Today’s report shows that there’s plenty to do moving forward, and I am looking forward to continuing our work to protect pollinators and the ecosystems that are dependent on them.”
“I’m excited that we have this report to help us understand the extent of pollinator decline in Colorado. While Colorado ranks fifth nationally for the rate of honey-bee die-offs, we haven’t known as much about native pollinating insects. This threatens our food production, biodiversity and health of our ecosystems,” said Rep. Cathy Kipp, D-Fort Collins. “We created the Native Pollinating Insects Protection Study in 2022 to identify ways to better protect and support the native Colorado pollinators, like different bee species, butterflies, and moths, that are also essential to maintaining a healthy environment. These findings will help direct future legislation to create a safer environment for our pollinators, protect our food supply and support biodiversity.”
Promoting policies that benefit native pollinating insects represents an opportunity to foster healthy and sustainable pollinator populations, especially in agricultural and urban habitats. This includes enhancing pollinator-friendly native plantings along I-76 following its designation as a Colorado Pollinator Highway by the Colorado Department of Transportation and other existing state plans such as the state’s Natural Areas Program, and Wildlife Action Plan among others.
“Working on the Pollinator Health Study has been an amazing opportunity to collaborate with so many locally, nationally, and internationally recognized experts in the field of pollinator conservation. In addition to the immense amount of information within the report, this study highlights the importance of collaboration between scientists and land management agencies to bring together the many facets needed for conserving native pollinating insects,” said Deryn Davidson, Sustainable Landscape State Specialist, Colorado State University Extension. “Having the existing research on Colorado pollinators paired with recommended land management practices in one, comprehensive document is an incredible tool for policy makers, land managers, and really anyone interested in actionable steps for pollinator conservation.”
Areas of immediate action and priorities highlighted by the Pollinator Report include:
- Priority 1: Protect imperiled native pollinating insects.
- Priority 2: Protect, restore, and connect pollinator habitats.
- Priority 3: Mitigate environmental changes that negatively impact pollinators and their habitats.
- Priority 4: Reduce the risks from pesticides to pollinating insects.
- Priority 5: Monitor and support native and managed pollinator health.
Governor Polis announced his annual budget proposal on November 1, focusing on ensuring Colorado is more affordable, sustainable, and liveable. The Governor included $100,000 to support education and incentives to encourage the use of pesticide alternatives in agricultural production and residential or commercial landscaping. On May 17, 2023, Governor Polis signed Neonic Pesticides as Limited-Use Pesticides, sponsored by Senators Kevin Priola and Sonya Jaquez Lewis and Representatives Kyle Brown and Cathy Kipp, which protects pollinators from harmful toxins.